
Suradnik:Imbehind/Evolucija
Evolucija (lat. evolutio: razvoj, razvitak) je znanstvena teorija iz područja biologije koja pretpostavlja da različite vrste biljaka, životinja i svih ostalih živih bića na Zemlji vuku podrijetlo iz već postojećih organizama, svojih predaka,[1] te da su sve uočljive morfološke i fiziološke razlike među njima nastale usljed niza manjih, često i neprimjetnih promjena tijekom mnoštva uzastopnih generacija.
Teorija evolucije je jedan od kamena temeljaca suvremene znanosti o biologiji, a zasniva se na opsežnim promatranjima, osobito iz područja paleontologije, embriologije, poredbene anatomije, poredbene fiziologije, genetike, molekularne biologije i behavioristike, znanstvene discipline koja proučava životinjsko ponašanje.[nedostaje izvor]
Živa bića evoluiraju kroz procese mutacije (slučajnih promjena na genima odnosno na DNK koja je nositelj nasljednih informacija), genetskog otklona (eng. Genetic drift) (slučajnih promjena u frekvenciji pojedinih genetskih varijacija unutar populacije tijekom vremena) i prirodne selekcije (ne-slučajnog i postepenog procesa kojim pojedine genetske varijacije i mutacije, a time i pripadajuće vidljive ili nevidljive osobine organizma, poput boje očiju, muskulature, gustoće krzna odnosno neke druge morfološke ili fiziološke osobine organizma, postaju manje ili više zastupljene u nekoj populaciji).[nedostaje izvor]
Većina genetskih mutacija niti pomažu niti odmažu individualnim pripadnicima neke populacije u borbi za preživljavanje. Kroz process genetskog otklona takvi mutirani geni se unutar populacije svrstavaju kao neutralni i mogu čistim slučajem opstati unutar populacije generacijama. Suprotno genetskom otklonu, prirodna selekcija nije slučajan process zato što djeluje na osobine organizma (genetske mutacije i varijacije) koje pozitivno ili negativno utječu na sposobnost preživljavanja i reprodukcije tog organizma u nekom okolišu.[2] Mutacija, genetski otklon i prirodna selekcija su kontinuirani i dinamički sastavni djelovi života kao pojave i tijekom vremena su oblikovali razgranatu strukturu drva života odnosno svo mnoštvo raznolikosti u sveukupnosti živog svijeta.
Uvod[uredi | uredi kôd]

Prema svim dosadašnjim znanstvenim opažanjima i spoznajama svi živi organizmi posjeduju nasljedni materijal u obliku gena, odnosno kodirajućih sekvencija DNK (u nekih virusa RNK), koje su naslijeđene od svojih roditelja i koje prosljeđuju svojim potomcima. Među potomcima postoje i genetske varijacije koje nastaju uslijed slučajnih promjena na lancu DNK, a koje nazivamo mutacijama, ili nastaju uslijed rekombinacije postojećih gena tijekom mejoze za vrijeme spolnog razmnožavanja.[3] Takve male razlike između roditelja i potomaka koje uvijek postoje su posve slučajne prirode. Ako su te promjene u odnosu na roditelje korisne, potomci imaju veću mogućnost preživljavanja, a time i reprodukcije. To znači da će veći broj potomaka u idućoj generaciji također posjedovati korisnu promjenu i time veću mogućnost preživljavanja i eventualnog razmnožavanja. Na ovaj način osobine koje doprinose tome da su organizmi bolje prilagođeni svojim životnim uvjetima postaju sve zastupljenije u populaciji kako se generacije izmjenjuju.[3] Takve malene genetske promjene mogu se vremenom akumulirati i rezultirati značajnijim promjenama pojedinih osobina unutar populacije sve do trenutka kada populacija, koja se sastoji od direktnih potomaka izvornih predaka, u odnosu na te iste pretke, predstavlja jednu posve novu živu vrstu. Može se prema tome zaključiti da je proces evolucije direktno odgovoran za mnoštvo različitih živih vrsta i oblika života na Zemlji[4].[5]

Uloga evolucije u nastanku novih vrsta je najvidljvija u slučajevima kada pojedina populacija postane izolirana od ostatka vrste. Do izolacije može doći iz jednostavnih razloga kao što su prirodna migracija populacije uslijed traženja novog životnog prostora ili prisilna migracija uzrokovana naglim kataklizmičkim promjenama (vulkanska aktivnost, potresi, požari, poplave), što uzrokuje geografsku udaljenost od ostatka vrste. Do izolacije populacije dolazi i uslijed drugih, postepenih prirodnih pojava kao što su otapanje ledenjaka i izmještanje toka rijeka ili djelovanja još sporijih ali posve neumoljivih prirodnih sila kao što je pomicanje tektonskih ploča koje uzrokuju stvaranje novih kontinenata i oceana, odnosno klimatskih promjena (pojava ledenih doba, nastanak pustinja). Značaj izolacije je u tome što fizička odvojenost pojedine populacije spriječava direktnu genetsku razmjenu sa ostatkom vrste pri čemu se i proces evolucije nastavlja odvijati posve odvojeno. Ukoliko je izolacija trajna, nakon dovoljno vremena i dovoljnog broja generacija, izolirana populacija može postati jedna sasvim nova vrsta[4].[5]

Iz temeljnih postavki teorije evolucije nužno proizlazi i teorija o univerzalnom zajedničkom porijeklu odnosno pretpostavlja postojanje prvotnog životnog oblika koji je predak svim danas poznatim živim vrstama.[6] Naime, ako se krene u rekonstrukciju evolucijske povijesti, generaciju za generacijom unatrag, broj živih vrsta se postepeno smanjuje kako se pojedine vrste kroz zajedničke pretke objedinjavaju u vrste koje su im prethodile, te se na kraju dolazi do jednog jedinog živog organizma koji je predak svih oblika života na zemlji koje danas poznajemo. Taj prvotni živi organizam iz kojeg su evoluirale sve poznate žive i izumrle vrste nazivamo posljednji univerzalni predak i smatra se da je živio u jednom od oceana koji su tada postojali negdje prije 3.8 milijardi godina.[7][8][9][10][11]
Iako Teorija evolucije ne objašnjava kako je život nastao, u potpunosti objašnjava svu golemu raznolikost života koju danas susrećemo i smatra se najvažnijom teorijom iz područja znanosti o životu na kojoj se temelje sva suvremena istraživanja i znanstvene spoznaje u biologiji. Teoriju evolucije kao i teoriju o univerzalnom zajedničkom porijeklu je prvi formulirao Charles Darwin 1869. u svojoj knjizi Postanak vrsta[12] te ga zbog toga danas smatramo ocem kako Teorije o evoluciji tako i moderne biologije.
Dokazi koji potvrđuju teoriju evolucije[uredi | uredi kôd]


Već su Darwin, ali i drugi biolozi 19. stoljeća pronašli uvjerljive dokaze o biološkoj evoluciji pomoću komparativnih studija živih organizama i njihove zemljopisne distribucije, ali i kroz proučavanje foslilnih ostataka izumrlih organizama.[13] Od Darwinovog vremena dokazi iz tih izvora su postali još jači i sveobuhvatniji, a nove discipline koje su se relativno nedavno pojavile - genetika, biokemija, fiziologija, ekologija, znanost o proučavanju ponašanja životinja (etologija) i posebice molekularna biologija - su isporučile snažne dodatne dokaze i detaljnu potvrdu ove znanstvene teorije. Dok je prikupljanje i izlaganje dokaza u Darwinovo vrijeme bio dugotrajan i mukotrpan postupak, a i sami dokazi su bili rijetki i teško dostupni, danas, koristeći tehnička dostignuća molekularne biologije, možemo direktno proučavati DNK i proteine bilo kojeg organizma što nam omogućava da smo u stanju rekonstruirati i najsitnije detalje evolucijske povijesti života. U 21. stoljeću jedino ograničenje u prikupljanju dodatnih direktnih dokaza u korist Teorije evolucije predstavlja problematično financiranje takvih istraživanja zbog činjenice da je znanstvena zajednica odavno prihvatila Teoriju evolucije kao znanstvenu činjenicu i ne smatra potrebnim daljnje rasipanje resursa sa ciljem dodatnog dokazivanja. Naime, biolozi koji proučavaju evoluciju više se ne bave prikupljanjem dokaza nego pokušavaju iz već prikupljenih dokaza stvoriti nove znanstvene spoznaje.
Fosilni zapisi[uredi | uredi kôd]
.jpg)
Kada živo biće ugine, njegovi ostaci obično bivaju uništeni ili od strane drugih živih bića ili od prirodnih procesa poput djelovanja vremena i erozije. U rijetkim slučajevima djelovi tijela - pogotovo tvrdi i otporni poput oklopa i ljuski, zubi ili kostiju - se mogu očuvati tako što igrom slučaja ostanu zakopani u blato, mulj ili pijesak, odnosno na neki drugi način budu zaštićeni od utjecaja grabežljivaca i prirodnih sila i vremena. Naposljetku, kroz procese petrifikacije i sedimentacije bivaju okamenjeni te postaju dijelom stijene koja je nastala uslijed djelovanja naknadno nataloženih slojeva tla.[14]

Paleontolozi su pronašli i proučavali fosilne ostatke tisuća živih bića koja su u prošlosti naseljavala zemaljske kontinente i oceane. Fosilni zapisi zorno prikazuju da su mnoge vrste izumrlih organizama po obliku prilično različite od onih koje susrećemo danas. Fosilni zapisi također pokazuju postepeni razvoj organizama i prelazak iz jednog oblika u drugi tijekom vremena koje se može egzaktno utvrditi uz pomoć metoda kronostratigrafije, pomoću kojih geološke slojeve i fosile koje u njima nalazimo stavljamo u precizan kronološki kontekst.[15] Primjerice, metodom radiometrijskog određivanja starosti stijena mjerimo količinu prirodnih radioaktivnih izotopa koji su preostali u nekom mineralu da bi pomoću njih utvrdili vrijeme koje je prošlo od njihova formiranja, a time i starost fosila koji se u tim stijenama nalaze.[16][17][18][19][20][21]

Radiometrijsko određivanje starosti stijena pokazuje da je Zemlja formirana prije otprilike 4.5 milijardi godina.[22] Najraniji fosili nalikuju na mikroorganizme poput bakterija i cijanobakterija (modro-zelene alge). Najstariji od tih fosila se pojavljuju u stijenama starim 3.5 milijardi godina u doba Prekambrija[10][11]. Najstariji poznati životinjski fosili, stari nekih 700 milijuna godina, iz geološkog razdoblja Ediakarija su fosili cjevastih i listolikih stvorenja mekih tijela.[23] Brojni fosili koji pripadaju i danas živućim koljenima sa zamjetnim mineralnim skeletom pojavljuju se u stijenama starim 540 milijuna godina u doba Kambrija. Ti organizmi su drugačiji od onih koje nalazimo danas i od onih koji su živjeli u međuvremenu. Neki od njih su radikalno drugačiji tako da su znanstvenici morali stvoriti novo taksonomsko koljeno kako bi ih mogli klasificirati. Prvi Kralježnjaci, životinje sa kralježnicom, pojavili su se prije 400 milijuna godina, a fosile prvih sisavaca nalazimo prije manje od 200 milijuna godina. Povijest života zabilježena fosilima u slojevima stijena predstavlja uvjerljiv dokaz za istinitost Teorije o evoluciji.

Fosilni zapisi nisu potpuni. Od vrlo malog broja organizama koji su očuvani u obliku fosila, još manji broj je pronađen, iskopan i proučavan od strane paleontologa. Međutim, u nekim slučajevima postepeni razvoj oblika tijekom vremena je detaljno rekonstruiran. Takav je slučaj sa evolucijom konja. Konja evolucijski, kroz fosilne zapise, možemo direktno povezati s životinjom veličine psa koja je imala nekoliko prstiju na svakoj nozi i zube prilagođene brštenju. Ta životinja, koju danas nazivamo prakonj (rod Hyracotherium), živjela je prije 50 milijuna godina. Suvremeni oblik, moderni konj (rod Equus), je puno veći, ima jedan prst na nozi kojeg zovemo kopito, a zubi su mu prilagođeni napasanju trave. Prijelazni oblici su jako dobro očuvani u obliku fosila, kao i mnoge druge vrste izumrlih konja koji su se razvijali u drugim smjerovima, ali u konačnici nisu ostavili živo potomstvo.[24]

Uz pomoć pronađenih fosila paleontolozi su rekonstruirali i radikalne slučajeve evolucijskih prijelaza u formi i funkciji. Primjerice, donja čeljust reptila odnosno gmazova sastoji se od nekoliko kostiju, a sisavci imaju samo jednu. Višak kostiju koje nalazimo u donjoj čeljusti reptila i dalje možemo naći u današnjih sisavaca, ali se ne nalaze više u čeljusti, već su to kosti srednjeg uha sisavaca i imaju funkciju u slušnom aparatusu tih životinja. Isprva, takva promjena se čini nevjerojatnom - teško je zamisliti kakvu bi funkciju, pogotovo korisnu, takve kosti imale tijekom prijelaznih oblika života između reptila i sisavaca. Međutim, paleontolozi su otkrili fosile čak dva prijelazna oblika gmazova koji naliku na sisavce, a koje zovemo terapsidi[25][26]. Ključna razlika u izgledu lubanje između terapsida i dotadašnjih gmazova bila je u funkcionalnoj vezi sa intenzivnijom obradom hrane, što znači da terapsidi više nisu samo gutali hranu, kako to rade gmazovi, već su je obrađivali čeljustima, što im je omogućavalo bolju i bržu obradu hrane. Naime, terapsidi su imali čeljust sastavljenu iz dva dijela što im je omogućavalo bolju pokretljivost čeljusti - u jednom dijelu se nalaze kosti koje nalazimo i u čeljusti sisavca, a u drugom nalazimo kosti koje će naposljetku postati čekić i nakovanj koje nalazimo u srednjem uhu suvremenih sisavaca. To je bio vrlo bitan uvjet za razvijanje aktivnog načina života koji je karakteristika sisavaca koje danas slobodno možemo smatrati jedinim preživjelim terapsidima.[27][28]


Za skeptične suvremenike Charlesa Darwina borbeni poklič je bila fraza "Karika koja nedostaje" koja označava tobože očigledan nedostatak fosilnih ostataka prijelaznih oblika između velikih čovjekolikih majmuna (hominidi) i čovjeka (homo) ili drugih kritičnih prijelaznih oblika, te je od tada u oporabi od strane neinformiranih i površnih osoba svih nacija i religija u smislu diskreditacije Teorije o evoluciji.[29] Međutim, pronađen je ne jedan, nego mnogi fosili prijelaznih oblika između različitih oblika života pa tako i prijelazni oblici između velikih čovjekolikih majmuna i ljudi koji zorno dokumentiraju evoluciju čovjeka.[30][31][32][33][34] Najstariji poznati fosili hominina, primata koji pripada ljudskoj lozi nakon što se odvojila od loze kojoj pripadaju veliki čovjekoliki majmuni, stari su 6-7 milijuna godina, dolaze iz Afrike i poznati su po imenima sahelantrop[35] i ororin[36] (pod zajedničkim imenom praeanthropus), a bili su predominantno bipedalni kad su se kretali po tlu i imali su vrlo mali mozak. Ardipitek[37] je živio prije 4.4. milijuna godina, također u Africi.
Brojni fosilni ostaci različitog afričkog porijekla poznati su pod zajedničkim imenom australopitek, koji je bio hominin koji se pojavio prije 3 do 4 milijuna godina. Australopiteci su bili uspravni, poput čovjeka, ali im je volumen lubanje bio mali - samo 500 cm3 (otprilike odgovara masi od 500g), što je usporedivo sa suvremenim gorilom ili čimpanzom, otprilike trećinom volumena mozga odraslog čovjeka. Australopitek je imao istovremeno karakteristike ljudi i majmuna - nisko čelo i izduženo majmunoliko lice, ali sa zubima u proporciji sličnoj onoj u ljudi.[38][39][40] Drugi rani hominini, koji su dijelom kontemporalni sa australopitecima uključuju kenyanthropusa i parantropa, stvorenja također relativno malih mozgova, iako su neke vrste parantropa imali veće tijelo. Parantrop danas predstavlja još samo postraničnu izumrlu lozu hominina.[41] Zajedno sa povećanjem volumena lubanje na više od 600cm3, u Homo habilisa, koji je živio prije 1,5 do 2 milijuna godina u Africi nalazimo sve više ljudskih karakteristika, kao i u Homo erectusa, koji je prije 0,5 do više od 1,5 milijuna godina tumarao područjima Afrike, Azije i Europe, a imao je volumen lubanje od 800 do 1000cm3 (težine mozga od 800 - 1100g).[42] Volumen mozga H. ergastera, H. antecesora i H. heidelbergensisa su bile usporedive sa volumenom lubanje H. erectusa i bili su djelomično suvremenici, iako su živjeli na različitim krajevima istočne hemisfere.
Evolucija čovjeka u fosilnim zapisima[uredi | uredi kôd]
Na donjoj galeriji možemo vidjeti, kroz prijelazne oblike, čitavu povijest razvoja čovjekovih predaka od najranijeg sahelantropa, kao prvog u nizu hominina koje više ne smatramo čovjekolikim majmunima do kromanjonaca, koje smatramo neposrednim predhodnicima ili čak podvrstom suvremene ljudske vrste Homo sapiens.
-
 Fosilizirana lubanja sahelnatropa
Fosilizirana lubanja sahelnatropa -
 Fosilizirana lubanja australopiteka
Fosilizirana lubanja australopiteka -
 Facijalna rekonstrukcija australopiteka
Facijalna rekonstrukcija australopiteka -
 Fosilizirana lubanja homo habilisa
Fosilizirana lubanja homo habilisa -
 Fosilizirana lubanja homo erectusa
Fosilizirana lubanja homo erectusa -
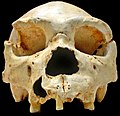 Fosilizirana lubanja homo heidelbergensisa
Fosilizirana lubanja homo heidelbergensisa -
 Fosilizirana lubanja homo rhodesiensisa
Fosilizirana lubanja homo rhodesiensisa -
 Lubanja kromanjonca
Lubanja kromanjonca -
 Digitalna facijalna rekonstrukcija kromanjonca
Digitalna facijalna rekonstrukcija kromanjonca -
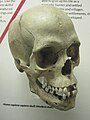 Lubanja modernog čovjeka
Lubanja modernog čovjeka -
 Portret žene masai ratnika
Portret žene masai ratnika









,_World_Museum_Liverpool.JPG)

Anatomske sličnosti i nesavršenost građe živih bića[uredi | uredi kôd]

Kosturi kornjača, konja, ljudi, ptica i šišmiša su na prvi pogled iznenađujuće slični, usprkos potpuno različitim načinima života i raznolikom okolišu u kojem te životinje žive. Neporeciva sličnost njihovih skeletalnih struktura, kad uspoređujemo broj i raspored pojedinih kostiju, može se lako uočiti ne samo na primjeru prednjih udova (ruke, noge, peraje, krila i sl.) već i u drugim dijelovima tijela.
Komparativna anatomija proučava podudarnosti ili naslijeđene sličnosti između organizama u građi kostiju ili među drugim djelovima tijela. Podudarnost u anatomskim strukturama je tipično vrlo velika između bliskih i srodnih organizama - primjerice između različitih vrsta ptica pjevica - ali postaje sve manja kako su organizami koje uspoređujemo sve manje srodni u odnosu na njihovu evolucijsku povijest. Može se uočiti da su sličnosti manje između n.pr. ptica i sisavaca nego među samim sisavcima, a još su manje između primjerice sisavaca i riba. Sličnosti u tjelesnoj građi nisu samo pojavna manifestacija evolucije nego pomažu i u rekonstrukciji filogenije evolucijske povijesti organizama.
Komparativna anatomija također otkriva zašto većina živih struktura nije savršena. Poput prednjih udova kornjača, konja, ljudi, ptica i šišmiša, ni drugi djelovi tjela nekog organizma nisu savršeno prilagođeni uporabnim uvjetima jer je njihov suvremeni oblik nastao modifikacijom postojeće, naslijeđene strukture, a nije bio namjenski projektiran, konstruiran, odnosno dizajniran ab ovo za točno određenu namjenu, bez ikakvih prethodnih referenci.
Primjerice, ako se razmotri struktura prednjih udova navedenih životinja, koja se sastoji od fundamentalno uvijek istih kostiju, sa manjim varijacijama u proporciji i dimenzijama, potpuno je neočekivano da reciklirajući jednaku anatomsku strukturu kornjače uspijevaju toliko dobro plivati, konj kasati, čovjek pisati, a ptica i šišmiš letjeti.
Ekstremni primjer anatomske nesavršenosti je i rekurentni laringealni živac ili povratni grkljanski živac (lat. nervus laryngeus recurrens, ), ogranak vagusa, koji u tijelu žirafe prevaljuje put od 4.6 metara od mozga niz vrat pa oko lijeve aorte i onda opet nazad do grkljana gdje ima funkciju upravljanja glasnicama.[43] Mozak i grkljan žirafe su inače udaljeni samo desetak centimetara, a razlozi ovakvom ekstremnom skretanju su čisto evolucijske naravi. Naime u ribolikih predaka današnjih kopnenih kralježnjaka (nadrazred tetrapoda) putanja živca bi bila direktna i vodila bi od mozga, prolazila neposredno uz srce pa ravno do škrga, kao što to je to slučaj u modernih riba. Međutim, tijekom evolucije, kako se vrat predaka žirafe sve više produljivao, a srce bilo smještano sve niže u tijelu tako se i povratni laringealni živac malo po malo produljivao kako bi se prilagodio toj značajnoj anatomskoj promjeni te je na kraju naprosto završio s "krive" strane srca, što je pak rezultiralo kružnom putanjom koju primjećujemo danas.[44]
U praktičnom smislu, dobar inženjer bi bio u stanju konstruirati bolje prilagođene udove za svaki navedeni primjer životinjskog kretanja ili lokomocije, odnosno predvidjeti izravniju, a time i smisleniju putanju rekurentnog laringealnog živca. Ali ako prihvatimo da su sve navedene životinje naslijedile svoju strukturu kostiju i druge anatomske značajke od zajedničkog pretka te da su poprimile današnji oblik isključivo tako da su se, podvrgnute procesu evolucije, postepeno prilagođavale različitom načinu života, tek tada evidentna sličnost njihovih anatomskih struktura ima smisla.
Ta nesavršenost anatomskih struktura živih organizama je uvjerljiv dokaz koji ide u prilog evoluciji, a suprotstavlja se antievolucijskim teorijama koje kao argument spominju takozvani inteligentni dizajn.
Embrionalni razvoj i zakržljali organi[uredi | uredi kôd]
Sam Darwin kao i njegovi sljedbenici su našli dokaze koji idu u prilog Teoriji evolucije i unutar embriologije, znanstvene discipline u čijem su fokusu živa bića u stadiju embrija, razvojnoj početnoj fazi života koja proučava razvoj organizama od oplođene jajne stanice do trenutka rođenja ili, u slučaju primitivnijih vrsta, dok se živo biće ne izleže iz oplođenog jaja. Svi kralježnjaci, od riba preko guštera do čovjeka, u ranom embrionalnom razvoju razvijaju se zapanjujuće slično, a onda, tijekom vremena, kako se embrio razvija i dozrijeva, zamjećuje se sve više razlika. Tih sličnosti između embrija ima više i traju dulje što su organizmi čiji se embriji proučavaju srodniji (na primjer embriji čovjeka i majmuna), a manje ako su organizmi različiti (na primjer embriji čovjeka i morskog psa). Može se zaključiti da zajednički obrasci razvoja reflektiraju evolucijsku bliskost. Embriji guštera i ljudi dijele razvojne obrasce naslijeđene od njihovog dalekog zajedničkog pretka - razvojni obrasci oba organizma su izmjenjeni samo utoliko što su se nakon razdvajanja do tada zajedničke loze sojevi guštera i ljudi razvijali u različitim smjerovima. Istovjetna faza razvoja embrija čovjeka i guštera reflektira ograničenja nametnuta naslijeđivanjem od istog zajedničkog pretka koje spriječava sve one promjene koje nisu uvjetovane njihovim različitim načinima života i drugačijim okolišem u kojem žive.
Na embrijima ljudi i drugih ne-vodenih kralježnjaka možemo primjetiti škržne otvore iako nikad neće razviti škrge i pomoću njih disati. Te otvore nalazimo na embrijima svih kralježnjaka iz razloga što dijele zajedničke pretke sa ribama kod čijih su se embrija te anatomske strukrure prvi put razvile. Ljudski embrij do četvrtog tjedna razvoja ima lijepo razvijen rep, koji maksimalnu duljinu postiže negdje oko šestog tjedna. Slične embrionske repove nalazimo i kod drugih sisavaca poput pasa, konja i majmuna, ali kod ljudi taj rep se postepeno skraćuje, sve do svog rudimentarnog oblika, trtične kosti, u kojem ga nalazimo u zdjelici potpuno razvijene osobe.
Bliske evolucijske veze između organizama koji na prvi pogled doživljavamo potpuno različitima mogu se ponekad razaznati po njihovim embrionalnim homologijama. Tako neke vrste priljepaka, inače sjedilačkih životinja, pripadaju koljenu Crustacea, što ih čini bliskim rođacima rakova, jastoga i škampa, što se nikako na prvi pogled ne bi moglo zaključiti budući da su spomenuti člankonošci pokretne, a ne sjedilačke životinje i naizgled nemaju nikakve veze sa spomenutim priljepcima. Međutim, u svom razvoju priljepci prolaze kroz stadij larve koji se zove nauplius u kojem slobodno plivaju morima i koji je bez ikakve sumnje veoma sličan ličinkama drugih rakova.
Rudimenti anatomskih cjelina vidljivi u embrionalnoj fazi, a koji se nikad u potpunosti ne razviju, poput škržnih otvora u ljudi, su uobičajeni kod svih vrsta životinja. Međutim, neki od njih, poput zakržljalog repa u ljudi, mogu zaostati i u zreloj fazi organizma kao podsjetnik na evolucijski proces i zajedničko porijeklo.
Najpoznatiji takav rudimentarni, zakržljali organ je slijepo crijevo ili crvuljak (lat. appendix vermiformis). Ovaj crvoliki organ je prikačen na kratki dio crijeva zvan cecum, koji se nalazi na mjestu spoja tankog i debelog crijeva. Ljudski crvuljak je oblikom zakržljao i njegova funkcija je vrlo mala ili nikakva, dok je u drugih životinja, poput zečeva i drugih biljojeda to potpuno razvijen organ sa značajnom probavnom funkcijom. Naime u njih veliki cecum i crvuljak pohranjuju biljnu celulozu kako bi omogućili njenu probavu uz pomoć bakterija. Osim crvuljka možemo navesti i druge primjere poput zakržljalih nogu koje nalazimo u zmija koje su nastale od guštera kao i u kitova koji su nastali od kopnenih sisavaca. U svakom slučaju, zakržljali organi su primjeri nesavršenosti koje vidimo u anatomskim strukturama mnogih organizama koji invalidiraju teorije kreacionizma i Inteligentnog dizajna, ali su pojave potpuno razumljive ako ih promatramo kao rezultate evolucije.
Geografska distribucija vrsta odnosno biogeografija[uredi | uredi kôd]

Darwin je vidio potvrdu svoje teorije o evoluciji i u zemljopisnoj rasprostranjenosti biljaka i životinja, a kasnije spoznaje su još više učvrstile njegova opažanja. Primjerice, na svijetu postoji oko 1500 poznatih vrsta vinske mušice (lat. Drosophila). Skoro jedna trećina od ukupnog broja žive na Havajima i nigdje drugdje, iako je ukupna površina arhipelaga manja od jedne dvadesetine površine Njemačke. Također, na Havajima postoji više od 1000 vrsta puževa i drugih kopnenih mekušaca koji ne žive nigdje drugdje. Ova neuobičajena raznovrsnost se može jednostavno objasniti evolucijom. Naime, Havajsko otočje je ekstremno izolirano i imalo je jako mali broj kolonizatorskih vrsta - biljaka i životinja koji su na Havaje dospjeli sa drugih mjesta na kojima su imale značajnije populacije. Kako na otočku prije njihovog dolaska nije bilo drugih kopnenih živih vrsta, budući da su Havaji vulkanskog porijekla i kao takvi su doslovno izronili iz mora u relatvno kratkom geološkom periodu, vrste koje su na Havaje dospjele na njima su naišle na veliki broj slobodnih ekoloških niša. U takvim izoliranim lokalnim ekosustavima te nove vrste su se mogle slobodno razvijati budući da nije bilo konkurentnih vrsta kao ni grabežljivaca koji bi ih spriječili da se množe. U takvim uvjetima došlo je do ubrzane diverzifikacije, procesa koji je omogućio da se popune sve dostupne ekološke niše, a kojeg nazivamo adaptivna radijacija.
.jpg)
Svaki od svjetskih kontinenata ima svoju zasebnu zbirku biljnog i životinjskog svijeta. Tako u Africi imamo nosoroge, nilske konje, lavove, hijene, zebre, žirafe, lemure, majmune sa uskim nosevima i repovima koje ne mogu omatati oko grana drveća, čimpanze i gorile. Južna Amerika, koja se proteže na istim zemljopisnim širinama kao i Afrika nema ni jednu od ovih životinja. Umjesto njih, u Južnoj Americi nalazimo pume, jaguare, tapire, ljame, rakune, oposume, armadile kao i majmune sa širokim nosevima i velikim repovima koje mogu omotati oko grana drveća po kojima se penju.
Ovakva biogeografska ćudljivost nije uzrokovana isključivo prilagođenošću životinja točno određenim životnim okolišima. Nema razloga da pomislimo da južno američke životinje nisu dobro prilagođene životu u Africi ili one afričke životu u Južnoj Americi. Havajsko otočje nije ništa pogodnije za život vinskih mušica od ostalih pacifičkih otoka, niti su manje gostoljubivi od drugih djelova svijeta za mnoge organizme koje na njima ne nalazimo. Zapravo, iako ni veliki sisavci nisu starosjedioci na Havajskom otočju, svinje i koze tamo uspješno preživljavaju, čak i kao posve divlje životinje, te se uspiješno množe sve od kada su ih na otok dovezli ljudi. Ovakva odsutnost mnogih živih vrsta iz gostoljubivog okoliša u kojem uspijevaju neobični varijeteti drugih vrsta može se lako objasniti evolucijom, koja pretpostavlja da vrste mogu postojati i evoluirati samo u zemljopisnim područjima koja su već kolonizirana od strane njihovih predaka.
Molekularna biologija[uredi | uredi kôd]
Molekularna biologija nam donosi do sada najdetaljnije i najuvjerljivije dokaze o ispravnosti Teorije o evoluciji. Kroz otkriće DNK i razumijevanje njene prirode, kao i razumijevanja prirode živih organizama na nivou enzima, bjelančevina i drugih molekula, molekularna biologija je pokazala da spomenute molekule sadržavaju informacije o porijeklu samih organizama kojih su integralni dio. Ovo je omogućilo znanstvenicima da rekonstruiraju događaje u evoluciji koji su prethodno bili nepoznati, kao i da potvrde ili prilagode saznanja o evoluciji koja su već bila poznata iz drugih znanstvenih disciplina. Preciznost rekonstrukcije evolucijskih događaja je ključni faktor koji dokaze iz polja molekularne biologije čini toliko značajnim. Drugi razlog je potvrda, na molekularnom nivou, Darwinove hipoteze o zajedničkom porijeklu svih živih organizama po kojoj su svi živi organizmi, od bakterije do čovjeka, povezani po podrijetlu od zajedničkog pretka.
U molekularnim komponentama živih organizama se susreće izražena uniformnost, kako u prirodi tih komponenata, tako i u načinu po kojem su te komponente sklopljene i iskorištene. U svim bakterijama, biljkama, životinjama, a time i ljudima, DNK se sastoji od različitih sekvenci ili slijedova uvijek istih četiriju nukleotida, a svi različiti proteini su sintetizirani od različitih kombinacija i sekvenci jednih te istih 20 amino kiselina, iako njih postoji nekoliko stotina. Genetski kod kojim je informacija sadržana unutar DNK u jezgri stanice, i kao takva se proslijeđuje proteinima, je praktično svuda potpuno podudarna. Slični metabolički putevi, sekvence biokemijskih reakcija, su korišteni od strane najrazličitijih organizama kako bi proizveli energiju i izgradili djelove živih stanica od kojih su građeni.
Ovakvo zajedništvo u istovjetnosti svih živih molekularnih struktura otkriva, i posve sigurno naznačava, genetski kontinuitet i jedinstveno porijeklo svih živih organizama. Ne postoji ni jedno drugo racionalno objašnjenje takve istovjetnosti baznih molekularnih struktura živih bića, kada postoje brojne druge alternativne strukture, koje su jednako vjerojatne, ali se uopće ne pojavljuju u živom svijetu. Genetski kod nam može poslužiti kao primjer. Svaka pojedina sekvenca od tri nukleotida u DNK jezgre djeluje kao šablona za proizvodnju uvjek jedne te iste amino kiseline u svim živim organizmima. Ovakva posljedična veza nije ništa nužnija nego što je u bilo kojem jeziku nužno da baš određena kombinacija slova predstavlja točno određeni predmet. Kad bi se utvrdilo da točno određene sekvence slova - npr. "planet", "žena", "drvo" - i u različitim knjigama imaju uvijek isto značenje, mogli bi biti sigurni da jezici na kojima su te knjige napisane, ako već nisu isti, sigurno imaju zajedničko podrijetlo.
Geni i proteini su dugačke molekule koje sadrže informaciju u sekvencama vlastitih komponenata na sličan način na koji rečenice hrvatskog jezika sadržavaju informaciju u sekvencama svojih slova i riječi. Sekvence koje tvore gene se prenose sa roditelja na potomstvo neizmjenjene, osim u rijetkim slučajevima kada se promjene dogode uslijed mutacija. Kao ilustraciju, možemo zamisliti da uspoređujemo sadržaj dvije knjige. Obje knjige imaju 200 stranica i sadrže isti broj poglavlja. Ako ih pobliže promotrimo, otrivamo da su knjige identične, od riječi do riječi, osim u rijetkim slučajevima, recimo jednom u 1000 riječi, kada je pojedina riječ drugačije napisana. U tom slučaju možemo zaključiti da je praktično nemoguće da su te dvije knjige neovisno napisane. Ili je jedna prepisana od druge ili su obje prepisane, direktno ili indirektno, od neke treće originalne knjige. Slično je i sa genetskim kodom. Ako svaki nukleotid u DNK gledamo kao jedno slovo, kompletna sekvenca nukleotida unutar DNK nekog složenijeg organizma bi ispunila nekoliko stotina knjiga sa stotinama stranica te sa nekoliko tisuća slova na svakoj stranici. Kada usporedimo takve "stranice", koje predstavljaju sekvence nukleotida, u tim "knjigama", koje predstavljaju različite žive organizme, uvelike istovjetan poredak slova na svakoj pojedinoj stranici pruža neoboriv dokaz o zajedničkom podrijetlu tih živih organizama.
Dva upravo predstavljana argumenta su zasnovana na različitim principima, iako oba govore u korist evolucije. Koristeći analogiju sa abecedom, prvim argumentom dokazujemo da jezici koji koriste isti rječnik, odnosno analogno, isti genetski kod i uvijek istih 20 amino kiselina, ne mogu biti neovisnog porijekla. Drugi argument koji opisuje sličnosti u sekvencama nukleotida unutar DNK, a time i sekvencama amino kiselina u proteinima, nam dokazuje da knjige sa vrlo sličim tekstom također ne mogu biti neovisnog porijekla.
Dokazi o evoluciji koji su otkriveni primjenom molekularne biologije idu i još dalje jer stupanj sličnosti u sekvencama nukleotida ili amino kiselina može biti iznimno precizno kvantificiran. Primjerice, u ljudi i čimpanza, molekula proteina citokroma C, koji služi vitalnoj funkciji staničnog disanja, sastoji se od uvijek istovjetno poredanog lanca od 104 amino kiseline. Međutim, razlikuje se od citokroma C u rezus majmuna u jednoj amino kiselini, od citokroma u konja u dodatnih 11 amino kiselina, a od onog u tune u dodatnoj 21. amino kiselini. Može se zaključiti da stupanj molekularne sličnosti DNK različitih organizama odražava vremensku i generacijsku udaljenost od zajedničkog pretka. Zbog toga se pretpostavke iz komparativne anatomije i drugih disciplina koje se bave evolucijom i poviješću evolucije mogu testirati pomoću ispitivanja DNK i proteina na molekularnom nivou ispitujući njihove sekvence nukleotida i amino kiselina.
Pouzdanost i autoritativnost ovakve vrste testova je neupitna. Svaki od tisuća gena i tisuća proteina sadržanih u organizmu omogućava neovisni test evolucijske povijesti tog organizma. Svi mogući testovi još uvijek nisu obavljeni, ali ih je obavljeno na tisuće i ni jedan od njih nije proizveo ni jedan jedini dokaz koji bi bio u suprotnosti sa Teorijom evolucije. Vjerojatno ni jedna druga pretpostavka iz bilo kojeg područja znanosti nije bila ovoliko detaljno testirana i temeljito potvrđena kao Teorija evolucije i zajedničkog podrijetla svih živih organizama.
Hijerarhijsko slaganje nasljednih osobina[uredi | uredi kôd]
Empirijska opažanja procesa evolucije[uredi | uredi kôd]
Sposobnost predviđanja ishoda empirijskih mjerenja[uredi | uredi kôd]
Povijesni razvoj Teorije o evoluciji[uredi | uredi kôd]
Rane ideje[uredi | uredi kôd]
U svakoj kulturi nailazimo na objašnjenje o postanku svijeta, živih stvorenja i ljudi. Tradicionalni judaizam i kršćanstvo objašnjavaju postanak živih bića i njihove prilagodbe okolišu poput krila, škrga, ruku i cvijetova kroz djelovanje svemogućeg Boga. Filozofi antičke Grčke imali su svoje mitove o postanku. Anaksimandar je tvrdio da se životinje mogu transformirati iz jedne vrste u drugu, a Empedoklo je špekulirao da su životinje sastavljene kombiniranjem već postojećih djelova.[45][46]
Bliži modernim idejama o evoluciji bili su stavovi ranih crkvenih otaca, kao što su Grgur Nazijanski i Augustin, koji su smatrali da nisu baš sve vrste životinja i biljaka stvorene od Boga, nego su se neke od njih razvile iz drugih božjih kreacija. Međutim, njihovi motivi nisu bili biološkog već religijskog karaktera - naime, smatrali su da bi bilo nemoguće smjestiti baš sve žive vrste na korablju koju je sagradio Noa, pa su zaključili da su neke vrste zasigurno nastale nakon potopa.
Ideja kako se živa bića mogu mijenjati uslijed prirodnih procesa od strane kršćanskih teologa srednjeg vijeka nije bila istraživana isključivo kao subjekt biologije, ali je, obično slučajno, ta mogućnost bila razmatrana od mnogih, uključujući Alberta Magnusa i njegovog učenika Tomu Akvinskog. Toma Akvinski je zaključio, nakon detaljnog razmatranja i rasprave u krugu svojih crkrvenih suradnika, da razvoj živih bića poput crva i muha iz nežive materije kao što je trulo meso nije inkompatibilan sa kršćanskom vjerom i folozofijom. Međutim, ostavio je drugima da odrede da li se tako stvarno i dogodilo.
Ideja napretka, a pogotovo vjerovanje u neograničen napredak ljudskog bića je bilo centralno vjerovanje prosvjetiteljstva 18. stoljeća, posebno u Francuskoj, među filozofima poput markiza Nicholasa de Condorceta i Denisa Diderota, kao i među znanstvenicima poput vojvode od Buffona, Georges-Louisa Leclerca. Međutim, vjera u napredak nije nužno vodila ka razvoju teorije o evoluciji. Pierre-Louis Moreau de Maupertuis je predložio pojam spontanog nastanka i izumiranja živih bića kao dio svoje teorije o postanku, ali nije iznio ništa slično teoriji evolucije, ideji transformacije vrsta iz jedne u drugu putem poznatih prirodnih procesa. Buffon, jedan od najvećih naturalista svog vremena je razmatrao i odbacio ideju mogućeg porijekla različitih vrsta od zajedničkog pretka. Njegov zaklučak je bio da kako novi organizmi nastaju spontano, iz organskih molekula, da može postojati onoliko životinja i biljaka koliko je mogućih spojeva tih istih organskih molekula.[47]
Engleski doktor Erasmus Darwin, djed Charlesa Darwina, u svojem djelu Zoonomija; Zakoni organskog života nudio je neke špekulacije povezane sa evolucijom, ali nisu bile naknadno razvijane te zbog toga nisu imale nikakav utjecaj na buduće teorije. Švedski botaničar Carolus Linnaeus, otac suvremene taksonomije, prvi je izradio hijerarhijski sistem klasifikacije biljaka i životinja koji se još uvijek koristi u moderniziranom obliku. Iako je osobno insistirao na nepromjenjivosti vrsta, njegov sistem klasifikacije je dao veliki doprinos prihvaćanju ideje o zajedničkom porijeklu vrsta.[48]
Veliki francuski naturalist Jean-Baptiste de Monet Lamarck, u svojim razmišljanjima držao se ideja prosvjetiteljstva kako živi organizmi utjelovljuju napredak sa čovjekom kao najsavršenijim primjerom takvog napretka. Iz te pozicije formulirao je na samom početku 19. stoljeća prvu jasnu teoriju evolucije. Prema njegovim shvaćanjima, organizmi su evoluirali kroz eone vremena iz nižih i jednostavnijih oblika u više i kompleksnije. Taj proces, kako je tvrdio, još uvijek traje i uvijek kulminira nastankom ljudskog bića. Kako se organizmi prilagođavaju svojem okolišu i stvaraju životne navike, nastaju modifikacije. Učestalo korištenje nekog organa ili anatomske strukture uzrokuje napredak i daljnji razvoj iste, dok zanemarivanje i nekorištenje dovodi do isčezavanja nepotrebnih anatomskih značajki.[49] Karakteristike stečene korištenjem ili nekorištenjem, prema njegovoj teoriji, se nasljeđuju. Ova njegova temeljna pretpostavka, kasnije nazvana nasljeđivanje stečenih karakteristika ili skraćeno Lamarkizam, je u potpunosti pobijena tijekom 20. stoljeća. Iako njegova teorija nije izdržala teret kasnijih znanstvenih spoznaja, Lamarck je načinio važan doprinos postepenom prihvaćanju biološke evolucije i stimulirao mnoga kasnija istraživanja.
Charles Darwin[uredi | uredi kôd]
Utemeljitelj moderne teorije o evoluciji je Charles Robert Darwin. Kako su njegov otac i djed bili doktori i on se upisao na studij medicine na sveučilištu u Edinburghu. Međutim, nakon dvije godine studiranja napustio je studije i otišao na sveučilište u Cambridgeu i pripremao se postati svećenik. Nije bio izvrsan student, ali je bio duboko zainteresiran za biologiju i povijest živog svijeta. 27. prosinca 1831., nekoliko mjeseci nakon što je diplomirao na Cambridgeu, u svojstvu prirodoslovca otplovio je brodom HMS Beagle na put oko svijeta koji je trajao do listopada 1836. Na tom putovanju Darwin je često bio u prilici izbivati s broda na dulje vrijeme i, za vrijeme takvih dugačkih izleta, prikupljati različite uzorke životinjskog i biljnog svijeta.
Otkriće fosilnih kostiju velikih izumrlih sisavaca u Argentini i proučavanje brojnih vrsta zeba na otočju Galapagos su događaji kojima se pripisuje Darwinov poseban interes za porijeklo vrsta. Godine 1859., nakon puno oklijevanja i nagovaranja od svog kolege prirodoslovca Alfreda Russela Wallacea, izdao je knjigu punog naziva "O postanku vrsta pomoću prirodnog odabira ili očuvanje boljih pasmina u borbi za opstanak", studiju koja je postavila teoriju evolucije i, što je još važnije, ulogu prirodnog odabira u određivanju tijeka evolutivnog procesa. Darwin je izdao i mnoge druge knjige sa temom evolucije, kao što je knjiga "Podrijetlo čovjeka i odabir ovisan o spolu", koja proširuje teoriju evolucije prirodnim odabirom na ljudsku evoluciju, ali njegovo najvažnije djelo, van svake sumnje, ostaje njegova revolucionarna prva knjiga koju danas skraćeno zovemo "Postanak vrsta".
Darwina su smatrali velikim intelektualnim revolucionarom koji je inaugurirao novu kulturnu eru čovječanstva, eru koja je bila druga i konačna faza kopernikanske revolucije koja je započela u 16. i 17. stoljeću pod vodstvom ljudi kao što su bili Nikola Kopernik, Galileo Galilei i Isaac Newton. Tada je kopernikanska revolucija označila početak doba moderne znanosti. Otkrića u astronomiji i fizici srušila su tradicionalne koncepte o svijetu i univerzumu u kojem živimo. Planet Zemlja više se nije smatrala centrom univerzuma, već je postala mali planet koji kruži oko jedne od mnoštva zvijezda. Godišnja doba i kiše koje omogućavaju proizvodnju hrane na usjevima, kao i razorne oluje i druge vremenske prilike i neprilike više nisu bile dio praznovjerja, već su postale poznate značajke prirodnih procesa, a kruženje planeta oko Sunca bilo je objašnjeno istim zakonima koji su se upotrebljavali za izračunavanje putanje topovskih kugli na Zemlji.
Značaj ovih i drugih sličnih otkrića je bio taj da su ona dovela do ideje o univerzumu kao materijalnom sustavu, koji je u neprestanom kretanju, a kojim vladaju isključivo prirodni zakoni. Funkcioniranje univerzuma više nije trebalo biti objašnjavano nepogrešivom voljom božanskog Tvorca. Umjesto toga, promišljanje o pojavama u prirodi prebačeno je u domenu znanosti gdje će one konačno biti objašnjene, čovjeku razumljivim, prirodnim zakonima. Prirodne pojave poput plime i oseke, pomrčine sunca i kretanja planeta na noćnom nebu mogle su se predvidjeti, kad god su uzroci tih pojava bili dovoljno poznati. Darwin je prikupio dokaze koji pokazuju da se evolucija dogodila, da različiti organizmi dijele zajedničko podrijetlo te da su se živa bića drastično mijenjala tijekom duge povijesti našeg planeta. Što je još važnije, znanstvenim stajalištem da je priroda sustav materije u gibanju kojim vladaju prirodni zakoni, jednim jedinim djelom - knjigom "Postanak vrsta", uspio je obuhvatiti i sav živi svijet.
Prije Darwina, postanak svih živih bića i njihova začuđujuća sposobnost prilagodbe, bili su pripisivani kreaciji sveznajućeg Boga. On je stvorio ribe u vodama, ptice u zraku i sve vrste kopnenih životinja i biljaka. Bog je svojim kreacijama podario škrge da mogu disati pod vodom, krila da mogu letjeti zrakom, oči da mogu vidjeti, a ptice i cvijeće tako je obojao kako bi ljudi mogli u njima uživati i prepoznati mudrost božju. Kršćanski teolozi, od Tome Akvinskog pa nadalje, tvrdili su da je prisustvo dizajna, namjere u stvaranju koja je vidljiva, štoviše očigledna kod svih živih bića, krunski dokaz postojanja vrhovnog Tvorca. Argument božanske namjere u kreaciji bio je "peti način" dokazivanja Boga u tezama Tome akvinskog.
Britanski teolog William Paley u svom djelu "Prirodna teologija" (1802.) poslužio se saznanjima o povijesti živog svijeta, fiziologijom i drugim tadašnjim znanstvenim spoznajama kako bi učvrstio tezu o tome kako je život stvoren od božanskog bića. Tako je ustvrdio da ako bi netko našao sat, čak i u nenaseljenoj pustinji, da bi sklad mnoštva djelova te naprave prisilio tu osobu da zaključi da je taj predmet napravio vješt urar. Nastavio bi analogijom sa ljudskim okom za koje je tvrdio da je puno složenije i savršenije od sata, ako se u obzir uzmu djelovi oka poput prozirne leće, mrežnice postavljene na točnoj udaljenosti kako bi se formirala slika i velikog živca koji prenosi signale do mozga. Ovaj argument u korist dizajna od strane stvoritelja je bio naizgled jak. Ljestve su napravljene za penjenje, nož za rezanje, a sat za određivanje vremena. Njihov funkcionalni dizajn nas upućuje na činjenicu da su te predmete izradili drvodjelac, kovač i urar. Slično tome, očigledno funkcionalan dizajn životinja i biljaka upućuju na postojanje božanskog Tvorca.
Darwinov genij je bio u tome što je uspio dati prirodno i znanstveno objašnjenje za organizaciju i funkcionalni dizajn živih bića. Prihvatio je činjenice prilagodbi - ruke su za hvatanje, oči za gledanje a pluća za disanje, ali je ipak uspio dokazati da svo mnoštvo biljaka i životinja, sa svim svojim prilagodbama i različitostima, može biti objašnjeno procesom prirodnog odabira, bez ikakve pomoći Tvorca ili nekog drugog natprirodnog mehanizma. To postignuće, kako će se pokazati, imalo je intelektualne i kulturne posljedice koje su bile dublje i trajnije od samih dokaza koje je Darwin upotrebio kako bi svoje suvremenike uvjerio u činjenicu o evoluciji.
Darwinova teorija prirodnog odabira je sažeta u samoj knjizi Postanak vrsta od samog Darwina:
Darwin je predložio prirodan odabir prvenstveno kako bi objasnio prilagodljivu prirodu živih bića jer je ispravno shvatio prirodan odabir kao proces koji aktivno promiče ili podržava prilagodbe. Evolucijske promjene tijekom vremena i evolucijska diverzifikacija (povećanje broja živih vrsta) nisu izravni proizvod prirodnog odabira, ali oni često uslijede kao njegov nusproizvod, jer prirodni odabir potiče prilagodbu različitim okruženjima.
Reakcije na Darwinovo djelo i promišljanja o evoluciji do kraja stoljeća[uredi | uredi kôd]
Izdavanje knjige "Postanak vrsta" je izazvalo veliko uzbuđenje u javnosti. Znanstvenici, političari, svećenici i drugi poznati ljudi svih zanimanja su raspravljali o Darwinovoj knjizi, braneći ili napadajući njegove ideje. Najistaknutiji sudionik kontroverznih rasprava neposredno nakon objave knjige bio je engleski biolog T. H. Huxley kojega su tada prozvali "Darwinov buldog". Huxley je žestoko branio teoriju evolucije u svojim artikuliranim, ali ponekad i zajedljivim javnim izlaganjima kao i u brojnim člancima koje je napisao u znak potpore Darwinu i njegovoj teoriji. Teorija evolucije prirodnim odabirom bila je omiljena tema društvenih događanja tijekom čitavih 1860-tih, pa i kasnije, ali su se oko nje povele i ozbiljne znanstvene rasprave i kontroverze, prvo u Britaniji, pa na kontinentu te naposlijetku i u SAD.
Suvremenik Darwina, engleski filozof sa velikim utjecajem u drugoj polovici 19. i početkom 20. stoljeća, bio je i Herbert Spencer (1820.–1903.). Spencer je prihvatio Darwinovu biološku teoriju i dao joj filozofski i društveni značaj. Postao je uporan promicatelj ideje evolucije koju je obilato koristio u svojim znanstvenim špekulacijama iz područja metafizike i društvenih znanosti. Njegove ideje su imale i određene negativne posljedice za ispravno shvaćanje i prihvaćanje Darwinove biološke teorije o evoluciji prirodnim odabirom. Osobno je popularizirao niz slogana poput onog najpoznatijeg "Najjači preživljavaju" ("Survival of the fittest"), kojeg je čak i Darwin preuzeo u kasnijim izdanjima Postanka vrsta. Usprkos tome Darwin je bio jako kritičan prema Spencerovom pristupu njegovoj teoriji pa je napisao:
Najopasniji elementi promišljanja Spencera i drugih srodnih filozofa su bile grube nadogradnje značaja Darwinovog izvorno biološkog termina "borba za preživljavanje" te primjene istih na područja ekonomije i društvenih znanosti, pristup koji danas nazivamo društvenim darvinizmom.
Najveća poteškoća s kojom se suočavala Darwinova evolucijska teorija bio je nedostatak adekvatne teorije nasljeđivanja, koja bi objasnila očuvanje varijacija na koje djeluje prirodni odabir tijekom niza generacija. Teorije o nasljeđivanju u Darwinovo vrijeme su držale da su karakteristike potomaka naprosto srednja vrijednost karakteristika njihovih roditelja, ali je i sam Darwin bio svjestan njihovih nedostataka - takve teorije nisu mogle objasniti očuvanje varijacija, zato što bi se, prema njima, razlike između potomaka prepolovile svakom novom generacijom, te bi se originalna varijacija brzo utopila u prosjeku već postojećih karakteristika. Međutim, "Kariku koja nedostaje" u Darwinovim argumentima vrlo brzo je objasnila Mendelova genetika. Otprilike u isto vrijeme kada je objavljen Postanak vrsta, redovnik Gregor Mendel je započeo opsežnu seriju eksperimenata na grašku u vrtu svog samostana u Brünnu, u Austro-Ugarskoj (današnje Brno, Češka Republika). Ti eksperimenti i analiza njihovih rezultata i danas se uče kao primjer primjene znanstvene metode. Mendelov članak, objavljen u časopisu prirodoslovnog društva u Brnu 1866, formulirao je fundamentalne principe teorije o nasljeđivanju koji su u primjeni do današnjih dana. Njegova teorija objašnjava biološko nasljeđivanje pomoću nasljednih faktora (koje danas zovemo genima) koje zasebno nasljeđujemo od oba roditelja, a koji se ne miješaju ili stapaju nego su strogo odijeljeni za vrijeme nastanka spolnih stanica, odnosno gameta. Mendelova otkrića su ostala nepoznanicom za Darwina, a i općenito nisu bila poznata do kraja stoljeća, kada su istovremeno ponovno otkrivena od strane brojnih znanstvenika na kontinentu te je tim otkrićem Darwinova teorija bila zaokružena.
U međuvremenu, darvinizmu se do kraja 19. stoljeća suprotstavljale alternativne teorije evolucije temeljene na ranijim teorijama. Naime, na temelju lamarkizma razvio se cijeli niz idealističkih mehanicističkih, tzv. neolamarkističkih hipoteza, koje se mogu svrstati u dvije skupine: psiholamarkizam i mehano-lamarkizam. Psiho-lamarkizam je utemeljio 1871. Edward Drinker Cope i po njemu je glavni pokretač evolucije psihička energija, koja se kod nižih životinja očituje u "nastojanju pod utjecajem nužde", kod viših životinja u "izboru", a kod razumnih životinja u "svjesnom izboru". Mehano-lamarkizam se razvio na osnovi nazora Herberta Spencera, koji je evoluciju organizama tumačio prijelazom tvari iz jednoličnog i indiferentnog stanja u raznoliko i diferencirano stanje. U skupinu mehano-lamarkističkih strujanja ulazi i teorija Theodora Eimera (1843.–1898.), poznata pod imenom Eimerova ortogeneza. Po toj je teoriji evolucijski proces koji ima sam po sebi točno određen smjer (tzv. usmjerena evolucija).
Osnovna su pitanja po kojima se darvinizam i lamarkizam razilaze pitanje nasljeđivanja stečenih svojstava i problem biološke svrhovitosti. Za lamarkiste je osnovno načelo evolucijskoga procesa neki teleološki postulat, "unutrašnja svrha", pa u svezi s time oni umanjuju ili sasvim ignoriraju ulogu prirodnog odabira, a to je po darvinizmu odlučujući faktor biološke evolucije.
Nasuprot različitim lamarkističkim smjerovima i hipotezama, koje podcjenjuju ili negiraju Darwinovo načelo prirodnog odabira, zoolog August Weismann ne priznaje nikakve druge faktore evolucije osim prirodnog odabira. Ta se teorija, nakon 1886. naziva neodarvinizam ili vajsmanizam. Weismann proširuje načelo prirodnog odabira i na procese koji se zbivaju u samom organizmu. On smatra da je nasljedna tvar ili zametna plazma sastavljena od niza nasljednih jedinica koje su okupljene u jezgri stanice. Najvažnije od njih bile bi posebne materijalne jedinice, "determinante", koje određuju svojstva pojedinih stanica. Determinante se mogu mijenjati, i to pojedinačno, neovisno o drugima, tako da cijeli organizam postaje mozaik pojedinih svojstava. Na taj način Weismann svu raznolikost evolucijskoga procesa svodi na kombinaciju nasljednih elemenata.
Weismannove determinante prihvatio je botaničar Hugo de Vries i postavio svoju mutacijsku teoriju evolucije. Po toj teoriji organizmi se mijenjaju samo u iznenadnim, skokovitim promjenama (mutacijama), a one se javljaju neovisno o utjecaju okoliša. Te mutacije nenadano proizvode nove vrste, koje su odmah stalne.[50] Na temelju novih znanstvenih činjenica Stephen Jay Gould i Niles Eldredge dali su nov model evolucijskih promjena koji su nazvali isprekidanom ravnotežom (engl. punctuated equilibria) ili punktualizmom, prema kojem se evolucijske promjene događaju naglo, nakon dugotrajna razdoblja stagnacije, za razliku od gradualizma – mišljenja da se evolucijski proces odvija postupno kroz dugo vrijeme.
Osim navedenih, postoji još cijeli niz evolucionističkih hipoteza i smjerova. Sva su te struje dale poneki doprinos razvoju biologije, ali kao evolucijske teorije imaju uglavnom povijesno značenje.
Moderna sintetska teorija evolucije[uredi | uredi kôd]
Ponovno otkrivanje Mendelove teorije nasljeđivanja 1900. godine od nizozemskog botaničara i genetičara Huga de Vriesa i drugih dovela je do naglašavanja mehanizma nasljeđivanja u evoluciji. De Vries je predložio novu teoriju evolucije poznatu pod imenom mutacionizam, koji u osnovi umanjuje značaj prirodne selekcije kao glavnog evolucijskog procesa. Prema De Vriesu (kojemu se pridužuju i drugi genetičari poput Williama Batesona iz Engleske) dvije vrste varijacija se odvijaju u organizmima. Prva je "obična" varijabilnost koju primjećujemo među individualnim pripadnicima vrste, a koja nema trejne posljedice na proces evolucije zbog toga što, prema De Vriesu, ne može "dovesti do prelaska granice jedne vrste, odnosno do postanka nove vrste, čak i u uvjetima najstrože i kontinuirane selekcije". Druga vrsta varijacija se sastoji od promjena izazvanih mutacijama, spontanim promjenama gena koje rezultiraju velikim promjenama organizma i naposlijetku nastanka nove vrste: "Nova vrsta zbog toga nastaje odjednom, proizvedena iz postojeće bez vidljive pripreme i bez prijelaznih oblika".
Mutacionizmu su se protivili mnogi znanstvenici, a posebno takozvani biometričari, predvođeni engleskim statističarem Karlom Pearsonom koji je branio Darvinov prirodni odabir kao glavni uzrok evolucije kroz kumulativni efekt malih, neprekidnih, individualnih varijacija (za koje su biometričari smatrali da se prenose iz generacije u generaciju bez ograničenja koja predviđaju Mendelovi zakoni nasljeđivanja).
Suprotstavljeni stavovi mutacionista (koje su u to vrijeme nazivali Medelijancima) i biometričara su pomireni u 20-tim i 30-tim godinama 20. stoljeća uslijed napretka na polju teorijske genetike. Naime, genetičari su pomoću matematičkih modela pokazali prvo, da se neprekinuti niz varijacija (na karakteristikama kao što su veličina tijela, broj izleženih jaja i slično) može objasniti Mendelovim zakonima i drugo, da prirodna selekcija koja djeluje kumulativno na malim varijacijama može dovesti do znatnih evolucijskih promjena u obliku i funkciji. Istaknuti članovi ove grupe teoretskih genetičara su R.A. Fisher and J.B.S. Haldane u engleskoj i Sewall Wright u SAD. Njihov rad je doprinio odbacivanju mutacionizma i, što je još važnije, postavio teoretski okvir za integraciju genetike u Darwinovu teoriju prirodne selekcije. Ipak, njihov rad je imao ograničen utjecaj na tadašnje biologe iz nekoliko razloga: bio je formuliran na matematičkom jeziku kojeg većina biologa nije razumjela; bio je skoro potpuno teorijski, sa malo empirijskih primjera i dokaza; i bio je ograničen u opsegu, većinom zanemarujući mnoge probleme poput specijacije (procesa nastanka novih vrsta), koji su bili od velike važnosti evolucionistima.
Veliki napredak je ostvaren 1937. sa izdanjem knjige "Genetika i postanak vrsta" Theodosiusa Dobzhanskog, američkog prirodoslovca i eksperimentalnog genetičara ruskog porijekla. Njegova knjiga je ponudila prilično opširno objašnjenje evolucijskih procesa kroz jezik genetičara i prizmu genetike kao znanosti, potpomognuto brojnim eksperimentalnim podacima i dokazima koji su podržavali njegove teorijske postulate. "Genetika i postanak vrsta" može se smatrati seminalnim djelom u formuliranju onoga što će kasnije postati poznato kao Sintetska teorija evolucije, koja je uspješno kombinirala Darwinovu prirodnu selekciju i Medelovu genetiku. Imala je enorman utjecaj na prirodoznance i eksperimentalne biologe, koji su vrlo brzo prigrlili novo razumijevanje procesa evolucije kao procesa genetskih promjena na pojedinim populacijama. Zanimanje znanstvenika za proučavanje procesa evolucije je nakon rada Dobzhanskog prilično naraslo pa su uskoro uslijedili drugi značajni doprinosi, proširujući sintezu genetike i prirodne selekcije na mnoštvo drugih znanstvenih polja unutar biologije kao znanosti.
Znanstvenici koje uz Dobzhanskog smatramo arhitektima sintetske teorije evolucije su američki zoolog njemačkog porijekla Ernst Mayr, engleski zoolog Julian Huxley, američki paleontolog George Gaylord Simpson i američki botaničar George Ledyard Stebbins. Ovi istraživači su uvelike doprinjeli procvatu instraživanja na polju evolucije unutar tradicionalnih biloških disciplina kao i unutar nekih novonastalih - posebno unutar populacijske genetike i kasnije evolucijske ekologije. Do 1950. prihvaćanje Darwinove teorije evolucije putem prirodne selekcije je bilo univerzalno među znanstvenicima, a sintetska teorija evolucije postala je općeprihvaćena.
Molekularna biologija[uredi | uredi kôd]
Fokus istraživanja nakon 1950. prebacio se na primjenu molekularne biologije na istraživanja na polju biološke evolucije. 1953. američki genetičar James Watson i britanski biofizičar Francis Crick su dešifrirali molekularnu strukturu DNK, nasljednog materijala sadržanog u kromosomima u jezgri svake stanice. Genetska informacija je kodirana unutar sekvence nukleotida koji čine lannce DNK molekula. Ova informacija određuje sekvence aminokiselina, građevnih jedinica molekula proteina, koji uključuju, među ostalim, strukturne proteine poput kolagena, respiratorne proteine poput hemoglobina, i brojne enzime odgovorne sa fundamentalne životne procese jednog organizma. Zbog toga, genetska informacija sadržana unutar DNK može se istraživati proučavanjem sekvenci amino kiselina u proteinima.
Sredinom 1960-tih, napredne, brze i jeftine laboratorijske tehnike i tehnologije poput elektroforeze i selektivne analize enzima su se počele primjenjivati na istraživanja razlika među enzimima i drugim proteinima. Primjena ovih tehnika na polju istraživanja evolucije omogućila je proučavanje problema koji se prije nisu mogli istraživati. Primjerice, istraživanje razmjera genetske varijacije u prirodnim populacijama (koji predstavljaju granice njihovog evolucijskog potencijala) i egzaktnog kvantificiranja promjene na genima prilikom postanka novih vrsta.
Usporedbe sekvenci aminokiselina unutar funkcionalno odgovarajućih proteina između različitih vrsta nam daju precizne, kvantitativne podatke o razlikama među tim vrstama nastalim od zajedničkih predaka. To je bio značajan napredak u odnosu na do tada tipično kvalitativne ocjene dobivene metodama komparativne anatomije i drugih tradicionalnih metoda proučavanja evolucije. 1968. japanski genetičar Motoo Kimura je predložio teoriju neutralnosti molekularne evolucije koja je predviđala da su, na nivou sekvenci nukleotida DNK i aminokiselina u proteinima, mnoge promjene neutralne te da nemaju skoro nikakvog utjecaja na funkciju molekula, a time i na prilagodbu organizma okolišu i njegovu sposobnost preživljavanja. Ispravnost te teorije ima za posljedicu i to da bi trebao postojati i evolucijski "molekularni sat". Drugim riječima, stupanj različitosti sekvenci aminokiselina ili nukleotida među pojedinim vrstama bi trebalo omogućiti pouzdanu procjenu vremena koje je proteklo od trenutka u kojem su se promatrane vrste spajale u jednu vrstu, zajedničkog pretka obaju promatranih vrsta. Ovo bi omogućilo rekonstrukciju evolucijske povijesti koja bi otkrila redoslijed grananja vrsta različitih sojeva i rodova poput onih koje su dovele do ljudi, čimpanzi i orangutana, kao i točnog vremena u prošlosti kada se jedna vrsta odcijepila od druge. Za vrijeme 70-tih i 80-tih godina 20. stoljeća, postalo je jasno da molekularni sat nije posve egzaktan. Svejedno, početkom 21. stoljeća ova teorija je nastavila davati najpouzdanije dokaze za potrebe rekonstrukcije evolucijske povijesti.
Laboratorijske tehnike DNK kloniranja i sekvenciranja omogućavanju nove načine istraživanja evolucije na molekularnom nivou. Plodovi rada sa ovim tehnologijama počeli su se akumulirati tijekom 1980-tih, prateći razvoj automatiziranih strojeva za DNK sekvenciranje i otkriće lančane reakcije polimeraze (PCR), jednostavne i jeftine tehnike koja u nekoliko sati može umnožiti milijarde ili triljune točno određenih DNK sekvenci ili gena za potrebe istraživanja. Veliki istraživački projekti poput Projekta ljudskog genoma su dodatno unaprijedili tehnologiju brzog i jeftinog dobivanja dugih DNK sekvenci. U prvih par godina 21. stoljeća, potpuni genomi su sekvencirani za više od 20 viših organizama, uključujući ljudska bića, kućne miševe (Mus musculus), štakore (Rattus norvegicus), vinske mušice (Drosophila melanogaster), komarce (Anopheles gambiae), obliće (Caenorhabditis elegans), parazite koji izazivaju malariju (Plasmodium falciparum), biljku gorušicu (Arabidopsis thaliana), i kvasac (Saccharomyces cerevisiae), kao i za mnoge druge mikroorganizme.
Dodatna istraživanja su provedena kako bi se istražili dodatni mehanizmi nasljeđivanja, uključujući epigenetsku modifikaciju (kemijsku modifikaciju specifičnih gena ili povezanih proteina), koja bi mogla objasniti sposobnost organizama da na potomstvo prenose značajke razvijene tijekom života jedinke.
Društveni i kulturni značaj teorije o evoluciji[uredi | uredi kôd]
Znanstveno prihvaćanje i proširenje na druge znanstvene discipline[uredi | uredi kôd]
Kritika i prihvaćanje od strane religija[uredi | uredi kôd]
Inteligentni dizajn i kritike[uredi | uredi kôd]
Koncepti i mehanizmi evolucije[uredi | uredi kôd]
Nasljeđivanje[uredi | uredi kôd]
Evolucija se u organizmima odvija kroz promjene u nasljednim osobinama - karakteristikama pojedinog organizma. U ljudi, na primjer, boja očiju je naslijeđena karakteristika i pojedinac može naslijediti nasljedno svojstvo smeđih očiju od jednog ili oba svoja roditelja.[51] Nasljedna svojstva kontroliraju geni, a kompletan skup gena u genomu jednog organizma se zove genotip.[52]
Kompletan skup vidljivih osobina koje čine strukturu i ponašanje organizma se zove fenotip. Ove osobine pojedinog organizma proizlaze iz interakcije genotipa s okolišem u kojem organizam živi.[53] Kao rezultat tog procesa, mnogi aspekti fenotipa jednog organizma nisu nasljedni. Na primjer, tamna koža nakon sunčanja u neke osobe nastaje interakcijom genotipa sa okolišem - sunčevim svjetlom. Stoga, tamna koža, sama po sebi, nije nasljedna osobina i djeca ljudi koji su boravili na suncu se ne rađaju sa brončanom bojom kože. Međutim, neki ljudi na suncu potamne lakše od drugih, zbog razlike u njihovom genotipu, a te su razlike, odnosno sam genotip, nasljedne. Također su poznati i primjeri ljudi sa naslijeđenom genetskom osobinom albinizma, koji uopće ne tamne i jako su osjetljivi na sunčevu svjetlost.[54]
Nasljedne osobine se proslijeđuju iz generacije u generaciju putem DNK, molekule koja nosi kodiranu genetsku informaciju.[52] DNK je dug polimer koji se sastoji od četri tipa baza. Redoslijed ili sekvenca baza duž jedne DNK molekule određuje genetsku informaciju, na sličan način na koji slova čine rečenicu. Prije nego što se stanica podijeli, DNK se kopira tako da obije novonastale stanice naslijeđuju istu DNK molekulu. Djelovi DNK molekule koji određuju pojedinačnu funkcionalnu cjelinu se zovu geni. Različiti geni imaju različite sekvence baza. Unutar stanica, duga vlakna DNK molekule tvore nakupine koje se zovu kromosomi. Specifične lokacije DNK sekvenci unutar kromosoma se zovu lokusi. Ako DNK sekvenca varira među pojedinim organizmima iste vrste, različiti oblici te sekvence se zovu alelama. DNK sekvence se mogu mijenjati zbog mutacija, što stvara nove alele. Ako se mutacija dogodi unutar gena, nova alela može utjecati na osobinu koju gen kontrolira, mijenjajući fenotip organizma.[55] Međutim, iako ova jednostavna veza između alela i nasljednih osobina postoji u nekim slučajevima, većina nasljednih osobina ima mnogo kompleksnije uzroke koji su kontrolirani interakcijom više gena.[56][57]
Nedavna istraživanja potvrđuju važne primjere nasljednih promjena koje ne mogu biti objašnjene sekvencama nukleotida unutar DNK organizma. Ove pojave se svrstavaju u skupinu epigenetskih sustava nasljeđivanja. [58] Metilacija DNK koja obilježava kromatin, samoodržive metaboličke petlje, utišavanje gena RNA interferencijom i trodimenzionalno usklađivanje strukture proteina (kao primjerice priona) su područja gdje su epigenetski sustavi nasljeđivanja otkriveni na nivou organizma.[59][60]
Sposobnost preživljavanja[uredi | uredi kôd]
Sposobnost preživljavanja je centralna ideja teorije evolucije. Može biti definirana u odnosu na genotip ili na fenotip koji djeluju u nekom prirodnom okolišu. U oba slučaja opisuje sposobnost organizma da preživi i da stvori potmstvo. Ona je jednaka je prosječnom doprinosu genetskom bazenu slijedeće generacije koja se sastoji od prosječne jedinke koja nosi točno određeni fenotip ili genotip koji je preživljavanju doprinio. Za ovu osobinu organizma se često koristi izraz Darwinova sposobnost preživljavanja, kako bi se razlikovala od fizičke sposobnosti preživljavanja jedinke.[61] Ako razlike među alelama nekog gena utječu na Darwinovu sposobnost preživljavanja, onda se frekvencija alela promijeniti tijekom više generacija. Alele sa većom sposobnošću preživljavanja tijekom vremena nalaze se sve češće. Taj proces se zove prirodan odabir.
Sposobnost preživljavanja individualnog organizma se manifestira kroz fenotip organizma. Na fenotip utječu kako okoliš u kojem se organizam razvio tako i geni, a sposobnost preživljavanja određenog fenotipa može biti različita u različitim okolišima. Stoga, sposobnost preživljavanja različitih jedinki sa istim genotipom nije nužno ista. Međutim, pošto je sposobnost preživljavanja pojedinog genotipa prosječna vrijednost, ona će reflektirati sposobnost stvaranja potomstva svih jedinki s istim genotipom u danom okolišu ili skupu različitih okoliša.
Inkluzivna sposobnost se razlikuje od pojedinačne sposobnosti preživljavanja tako što uključuje sposobnost alele u jednoj jedinci da potpomaže preživljavanje i stvaranje potomstva drugih jedinki s kojima dijeli istu alelu puno prije nego drugim jedinkama koje imaju neku drugu alelu. Jedan od mehanizama inkluzivne sposobnosti preživljavanja je odabir po srodstvu.
Sposobnost preživljavanja se često definira kao statistička vrijednost, nasuprot mjerenja stvarnog broja potomstva. Primjerice, prema Maynard Smithu, "Sposobnost preživljavanja je svojstvo ne jedinke, nego skupine jedinki. Zbog toga izraz 'očekivani broj potomstva' znači prosječan broj, a ne stvaran broj potomstva neke jedinke. Ako bi prvi čovjek sa genom koji bi mu omogućavao levitaciju bio pogođen munjom dok je ležao u koljevci, to ne bi dokazalo da novi genotip ima nisku sposobnost preživljavanja, već da baš to dijete nije imalo sreće".[62]
Varijacije[uredi | uredi kôd]


- Za daljnje informacije vidi gensku raznolikost i populacijsku genetiku.
Fenotip individualna organizma rezultat je kako njegova genotipa tako i utjecaja okoliša u kojem živi. Znatan dio varijacije u fenotipovima u nekoj populaciji uzrokovan je razlikama u njezinim genotipovima.[57] Moderna evolucijska sinteza definira evoluciju kao promjenu te genske varijacije tijekom vremena. Frekvencija nekog pojedinačnog alela postat će više ili manje prevalentna u odnosu na druge forme tog gena. Varijacija iščezava kada nov alel dosegne točku fiksacije – kada on ili iščezne iz populacije ili u potpunosti nadomjesti predački alel.[63]
Prirodna će selekcija jedino uzrokovati evoluciju ako postoji dovoljno genske varijacije u populaciji. Prije otkrića mendelovske genetike uobičajena je hipoteza bilo miješano nasljeđivanje. No s miješanim nasljeđivanjem genska bi varijanca ubrzo nestala što bi evoluciju prirodnom selekcijom učinilo nevjerojatnom. Hardy-Weinbergov princip pruža rješenje načina na koji se varijacija održava u populaciji Mendelovim nasljeđivanjem. Frekvencije alela (varijacije u genu) ostat će konstantne u odsustvu selekcije, mutacije, migracije i genskog drifta.[64]
Varijacija nastaje zbog mutacija genskog materijala, reorganizacije gena tijekom spolnog razmnožavanja i migracije između populacija (genski tok). Unatoč konstantnu uvođenju nove varijacije zbog mutacije i genskog toka, većina je genoma neke vrste identična u svim jedinkama te vrste.[65] No čak i relativno male razlike u genotipu mogu dovesti do dramatičnih razlika u fenotipu: na primjer, čimpanze i ljudi se razlikuju samo u oko 5% svojeg genoma.[66]
Mutacija[uredi | uredi kôd]
- Za daljnje informacije vidi mutaciju.

Mutacije su promjene u sekvenciji DNA staničnog genoma. Kada se mutacije dogode, one mogu ili biti bez ikakva efekta izmjenjujući proizvod nekog gena ili spriječiti gen da funkcionira. Na osnovi proučavanja mušice Drosophila melanogaster pretpostavlja se da ako mutacija promijeni protein koji gen proizvodi, to će vjerojatno biti štetno, oko 70% takvih mutacija imat će štetne efekte, a preostale će biti ili neutralne ili slabo povoljne.[67]
Mutacije mogu obuhvaćati velike djelove kromosoma tako da budu duplicirani (obično genskom rekombinacijom), koja može uvesti dodatne kopije gena u genom.[68] Dodatne kopije gena su glavni izvor sirovine potrebne da bi novi geni evoluirali.[69] Ovo je veoma važno budući da većina novih gena evoluira unutar familija gena od već postojećih gena koji dijele zajedničke pretke.[70] Na primjer, ljudsko oko koristi četiri gena kako bi stvorile strukture koje su osjetljive na svjetlo: tri za razlikovanje boja i jednu za gledanje u slabim svjetlosnim uvjetima. Sva četiri su nastala od jednog gena koji im je bio predak.[71]
Novi geni mogu biti stvoreni od gena pretka kada duplicirana kopija mutira i stekne novu funkciju. Ovaj proces je mnogo jednostavniji jednom kada je gen dupliciran zato što povećava redundanciju sistema. Jedan gen u paru istovjetnih gena može mutirati i time steći novu funkciju, dok drugi nastavlja izvršavati svoju originalnu funkciju.[72][73] Drugi oblici mutacija mogu čak stvoriti potpuno nove gene od prethodno nekodirajuće DNK.[74][75]
Generacija novih gena može također uključivati umnožene djeliće postojećih nekoliko gena pri čemu se ti umnoženi fragmenti rekombiniraju u nove kombinacije sa novom funkcionalnošću.[76][77] Kada su novi geni sklopljeni mješanjem prethodno postojećih djelova, domene proteina djeluju kao moduli sa jednostavnim neovisnim funkijama koje se mogu miješati kako bi se proizvele nove kombinacije sa novim i kompleksnim funkcijama.[78] Primjerice, poliketidne sintaze su veliki enzimi koji čine antibiotike. Oni sadrže i do stotinu neovisnih proteinskih domena od kojih svaka katalizira jedan korak u cjelokupnom procesu, poput diskretne operacije na pokretnoj traci.[79]
Spol i rekombinacija[uredi | uredi kôd]
- Za daljnje informacije vidi spolno razmnožavanje, gensku rekombinaciju i evoluciju spolnog razmnožavanja.
U bespolnih organizama, svi geni su naslijeđeni zajedno odnosno geni su vezani, jer se ne mogu miješati s genima drugih organizama za vrijeme reprodukcije. Nasuprot tome, potomci organizama koji imaju spol sadrže nasumične mješavine kromosoma njihovih roditelja koji su nastali kao proizvod neovisnog slaganja. U sličnom procesu nazvanom homologna rekombinacija, seksualni organizmi razmjenjuju DNK između dvaju odgovarajućih kromosoma.[80] Rekombinacija i preslagivanje ne mijenjaju frekvencije alela nego umjesto toga mijenjaju povezanost istih, rezultirajući potomcima sa novim kombinacijama alela.[81] Spol obično povećava genetsku varijaciju i može ubrzati tijek evolucijskog procesa.[82][83]
Genski tok[uredi | uredi kôd]
- Za daljnje informacije vidi genski tok.
Genski tok je razmjena gena među populacijama i među vrstama.[84] Zbog toga može biti izvor varijacije koja je nova u populaciji ili vrsti. Genski tok može biti uzrokovan kretanjem individualnih jedinki među različitim odvojenim populacijama organizama, kao na primjer kretanje pojedinog miša među kopnenim i obalnim populacijama ili kretanje polena trave između populacija sa visokom tolerancijom na teške metale i visokom osjetljivošću na teške metale.
Transfer gena između vrsta uključuje formiranje hibridnih organizama i horizontalni genski transfer. Horizontalni genski trasfer je transfer genetskog materijala od jednog organizma do drugog, koji prvom nije potomak. Ova pojava je uobičajena među bakterijama. [85] U medicini, ova pojava doprinosi širenju otpornosti na antibiotike tako da jedna bakterija, koja razvije gene koji joj daju otpornost, može te iste gene rapidno prenijeti drugim vrstama. [86] Zabilježen je i horizontalni transfer gena od bakterija prema eukariotima kao što je kvasac Saccharomyces cerevisiae i buba azuki graha Callosobruchus chinensis.'[87][88] Primjer transfera na većoj skali su eukariotski bdeloidni kolnjaci, koji su primili čitavu skupinu gena od bakterija, gljiva i biljaka.[89] Virusi također mogu prenijeti DNK između organizama, omogućavajući transfer gena čak i preko granica bioloških carstava.[90]
Prijenos gena na većoj skali se također dogodio između predaka eukariotskih stanica i bakterija za vrijeme stjecanja kloroplasta i mitohondrija. Moguće je da su eukarioti i nastali horizontalnim transferom gena između bakterija i archaea.[91]
Mehanizmi evolucije[uredi | uredi kôd]
Prirodna selekcija[uredi | uredi kôd]
Prirodni odabir je neslučajni proces kojim određene biološke značajke odnosno genotipovi nestaju iz populacije tijekom vremena kao posljedica slabije prilagođenosti uvjetima života nositelja te ili tih značajki. Kao otkriće, načelo odabira glavni je Darwinov doprinos teoriji evolucije, koju on objašnjava borbom za opstanak među pojedinim organizmima, od kojih preživljavaju najsposobniji. Prirodni odabir, dakle, Darwin primjenjuje na jedinke a ne na populacije, a okoliš kao selektivna sila odabire varijante koje su joj najbolje prilagođene. Darwin je time naglasio negativno gledište selekcije, ističući da je ona neumoljiva snaga koja uništava jedne a unaprjeđuje druge.
Usmjerena mutacija[uredi | uredi kôd]
Genetski drift[uredi | uredi kôd]
Genetsko stopiranje[uredi | uredi kôd]
Genetski tok[uredi | uredi kôd]
Rezultati evolucijskog procesa[uredi | uredi kôd]
Adaptacija[uredi | uredi kôd]
Usporedna evolucija ili koevolucija[uredi | uredi kôd]
Kooperacija[uredi | uredi kôd]
Postanak novih vrsta ili specijacija[uredi | uredi kôd]
Izumiranje[uredi | uredi kôd]
Podjela evolucijskih procesa[uredi | uredi kôd]
S obzirom na opseg i određene karakteristike evolucijskih procesa, moguće je razlučiti
- mikroevoluciju,
- makroevoluciju i
- megaevoluciju kao stupnjeve ili razine u neprekidnom evolucijskom nizu.[92]
Mikroevolucija[uredi | uredi kôd]
Mikroevolucija sadrži manje sukcesivne promjene u genskoj zalihi određene populacije od jedne do druge generacije.[93] U njoj djeluju osnovne sile evolucije. Procesi mikroevolucije vode stvaranju novih vrsta, odnosno populacija. Specijacija je nastanak novih vrsta. Naziva se i populacijska divergencija. Cijeli tijek evolucije ovisi o podrijetlu novih populacija koje imaju adaptivnu sposobnost veću od svojih predaka. U nastanku novih vrsta uz osnovne sile djelatne su i dodatne sile, koje djeluju dulje vrijeme. Glavni je faktor izolacija. Ima nekoliko oblika izolacije: geografska, ekološka, reprodukcijska, genska. U geografskoj izolaciji proces se odvija tako da se početna populacija širi od sredine svojega prebivališta u susjedna područja. U početku se izmjena gena nastavlja između svih dijelova populacije. Postupno se male populacije počinju fragmentirati i ustaljivati u najpogodnijim prebivalištima, pa jedne od drugih postaju odijeljene ekološki nepovoljnim područjima. Nakon određenog vremena izolacije započinje fragmentacija u još manje populacije ili demove. Dijeljenje početne populacije u homogene serije dema poznato je kao ponašanje svih živućih organizama. Svaki dem zadržava isti genski sastav kakav je imala populacija predaka, ali su ekološke barijere genskoj izmjeni između demova temelj genske divergencije između njih. Svaki izolirani dem biva od početka modificiran silama mikroevolucije. Nakon nekog vremena demovi se počnu međusobno razlikovati, iako još uvijek ostaju otvoreni genskoj izmjeni. Primarni je način postanka vrste alopatrijska specijacija; u njoj prostorna izolacija, pojačana geografskim ili ekološkom barijerom, djeluje tako da dvije prvotno vrlo slične populacije postaju genski izolirane. Ekološka izolacija sastoji se u tom da populacije koje žive na istom geografskom području postaju izolirane faktorima okoliša tako da zauzmu odijeljene biotope ili nastanjuju specijalne ekološke niše. Taj proces vodi stjecanju specifično usklađenih gena i gubljenju prvotno zajedničkih oznaka, a to unapređuje fiziološku izolaciju. Dogodi li se da se takve populacije, nakon dugog vremena, ponovno nađu zajedno, unatoč činjenici da nastanjuju isti areal, one se više međusobno ne križaju. Reprodukcijska izolacija sastoji se u razlikama u ponašanju u doba parenja (npr. u ptica razlike u glasanju), u razlikama s obzirom na razdoblje i sezonu parenja, u morfološkoj neusklađenosti kopulacijskih organa što priječi nesmetano neograničeno križanje (panmiksiju). Čini se da geografsko odjeljivanje odlučno utječe na mehanizam te izolacije. Genetska izolacija nastupa kao posljedica promjena u kromosomskoj garnituri ili u poretku gena. Kada nastupe takve promjene, kromosomi dotičnih vrsta ne mogu se spojiti, pa više-manje nepravilno djeluju ako i dođu u dodir. Zbog antagonističnosti genetskoga sastava roditeljskih vrsta, gamete ili ugibaju već u samom početku procesa oplodnje ili pak, ako se nastavi razvoj zametka, kote se životno nesposobni ili sterilni hibridi. Nastanak novih vrsta kroz samu genetsku izolaciju rijedak je, ali zajedno s geografskom izolacijom dobiva veliko značenje.
Makroevolucija[uredi | uredi kôd]
Makroevolucija ili adaptivna radijacija evolucija je iznad razine vrste, a karakterizira ju dijeljenje ili fragmentacija, koja dovodi do postanka i razvoja većeg broja adaptivnih tipova. Dok u mikroevoluciji i specijaciji nastaju posebne adaptacije, u makroevoluciji razvoj se odvija od opće adaptacije divergentno prema mnogobrojnim posebnim adaptacijama. Makroevolucija obuhvaća razvoj viših sistematskih skupina (porodica, red, razred).[94][95][96] Nju karakterizira:
- dioba skupina u mnogo novih podskupina;
- prodor u nove sredine;
- organiziranje različitih struktura i načina života.
Osnovna je karakteristika evolucije iznad razine vrste pomicanje skupine organizama u nove adaptivne zone. Kako bi organizam mogao proći kroz zonu ekološke nestabilnosti u novu adaptivnu zonu, on mora imati otvoren evolucijski i ekološki pristup u nju, tj. skupina mora već imati neka prilagodljiva svojstva, a zona ne smije u sebi imati jakog konkurenta. Prvi prodor u novu zonu zahtijeva samo opću adaptaciju; poslije evolucija vodi specijaciji. Važno je svojstvo makroevolucije također usmjeravanje razvoja nekih svojstava prema samo jednoj razvojnoj liniji. Evolucija gmazova paleontološkim je nalazima najbolje dokumentiran primjer makroevolucije. Čini se da su određene karakteristike adaptivne radijacije, utvrđene u evoluciji gmazova, osnovne značajke makroevolucije u svim skupinama, a to su:
- Do makroevolucije dolazi uvijek nakon stjecanja nove opće adaptacije ili nakon ulaska u novu adaptivnu zonu (kod gmazova je nastala opća adaptacija za život na kopnu).
- Makroevolucija nije linearna već zrakasta. Radijacija se nastavlja na opću adaptaciju i na ulaz u novu adaptivnu zonu time što se razvija specijalna adaptacija u različitim divergentnim novim linijama. (Tipični primjer radijacija arkosaura od općeg oblika tekodonta .)
- Adaptivna radijacija stvara evolucijske linije koje, s obzirom na specijalne adaptacije, konvergiraju prema drugim sistematski udaljenim skupinama, a koje se razlikuju po načinu opće adaptacije. (Ihtiosauri su primjer evolucijske konvergencije prema ribama.)
- Makroevolucija stvara skupine paralelnih specijalnih adaptacija između divergentnih ali srodnih linija, koje imaju zajedničku osnovu opće adaptacije. (Fitosauri i krokodili iz trijasa danas su primjer takva paralelizma i ekološke zamjene.)
- U makroevoluciji zbiva se definitivno propadanje skupina organizama. Kako je opća adaptacija zamijenjena specijalnom, skupine se kruto specijaliziraju na uske adaptivne podzone i izgube sposobnost za pomak u nove veće zone. Budući da se sve adaptivne zone moraju konačno promijeniti, skupine zatvorene u usku zonu moraju propasti. (Primjer su za to Rhynchocephalia, iznimno uspješan red sve do krede, od kojega živi još samo jedan rod (premosnici) sa samo dvije vrste u Novom Zelandu.)
Megaevolucija[uredi | uredi kôd]
Megaevolucija je pojava novih bioloških sustava ili tipova organizacije. Kroz tri milijarde god. razvilo se gotovo 200 takvih tipova. Njihovo podrijetlo najvažniji je događaj u evoluciji, vrlo kompliciran i najmanje istražen. Megaevoluciju karakteriziraju: 1. pokušaji i istraživanja raznosmjernih linija od polaznoga pretka da prijeđu ekološku granicu i uđu u novu zonu, dok jednoj konačno to i ne uspije; 2. prodor i promjena uvijek su brzi; 3. nova zona uvijek je ekološki pristupačna i nema suparnika, a zahtijeva novi općeadaptivni tip za invaziju (npr. razvoj gmazova od vodozemaca).
Glavno je svojstvo organske evolucije divergencija koju usmjeruje prirodna selekcija kao glavna sila. Na svim razinama evolucije djelatne su osnovne sile, ali se javljaju i sve složenija djelovanja manje poznatih sila.
Evolucijska povijest života[uredi | uredi kôd]
Nastanak života[uredi | uredi kôd]
Zajedničko porijeklo[uredi | uredi kôd]
Evolucija života[uredi | uredi kôd]
Molekularna evolucija[uredi | uredi kôd]
Bla Bla

[[kategorija:genetika]] [[kategorija:evolucijska biologija]]
Izvori[uredi | uredi kôd]
- ↑ Hall, B. K.; Hallgrímsson, B., ur. 2008. Strickberger's Evolution četvrto izdanje izdanje. izdavač: Jones & Bartlett. ISBN 0-7637-0066-5
- ↑ Garvin-Doxas, K.; Klymkowsky, M. W. 2008. Understanding Randomness and its Impact on Student Learning: Lessons Learned from Building the Biology Concept Inventory (BCI). CBE Life Sciences Education. 7 (2): 227–233. doi:10.1187/cbe.07-08-0063. PMC 2424310. PMID 18519614
- ↑ a b Gregory, T. R. 2009. Understanding Natural Selection: Essential Concepts and Common Misconceptions. Evolution: Education and Outreach. 2 (2): 156–175. doi:10.1007/s12052-009-0128-1
- ↑ a b Understanding Evolution: your one-stop source for information on evolution (web resource). The University of California Museum of Paleontology, Berkeley. 2008. Pristupljeno 23. siječnja 2008. journal zahtijeva
|journal=(pomoć);|contribution=zanemaren (pomoć) - ↑ a b Cavalier-Smith T. 2006. Cell evolution and Earth history: stasis and revolution. Philosophical Transactions of the Royal Society B. 361 (1470): 969–1006. doi:10.1098/rstb.2006.1842. PMC 1578732. PMID 16754610
- ↑ Theobald, D. L.I. 2010. A formal test of the theory of universal common ancestry. Nature. 465 (7295): 219–22. Bibcode:2010Natur.465..219T. doi:10.1038/nature09014. PMID 20463738
- ↑ Doolittle, W. F. 2000. Uprooting the tree of life (PDF). Scientific American. 282 (6): 90–95. doi:10.1038/scientificamerican0200-90. PMID 10710791.
- ↑ Glansdorff, N.; Xu, Y; Labedan, B. 2008. The Last Universal Common Ancestor: Emergence, constitution and genetic legacy of an elusive forerunner. Biology Direct. 3: 29. doi:10.1186/1745-6150-3-29. PMC 2478661. PMID 18613974.
- ↑ Yoko Ohtomo, Takeshi Kakegawa, Akizumi Ishida, Toshiro Nagase, Minik T. Rosing. 8. prosinca 2013. Evidence for biogenic graphite in early Archaean Isua metasedimentary rocks. Nature Geoscience. doi:10.1038/ngeo2025. Pristupljeno 9. prosinca 2013.CS1 održavanje: upotreba parametra authors (link)
- ↑ a b Borenstein, Seth. 13. studenoga 2013. Oldest fossil found: Meet your microbial mom. AP News. Pristupljeno 15. studenoga 2013.
- ↑ a b Noffke, Nora; Christian, Daniel; Wacey, David; Hazen, Robert M. 8. studenoga 2013. Microbially Induced Sedimentary Structures Recording an Ancient Ecosystem in the ca. 3.48 Billion-Year-Old Dresser Formation, Pilbara, Western Australia. Astrobiology (journal). doi:10.1089/ast.2013.1030. Pristupljeno 15. studenoga 2013.
- ↑ Darwin, C. 1859. The Origin of Species by Means of Natural Selection. John Murray. str. 490
- ↑ The Fossil Record - Life's Epic. The Virtual Fossil Museum. Pristupljeno 31. kolovoza 2007.
- ↑ McGraw-Hill Encyclopedia of Science & Technology. McGraw-Hill. 2002. str. 58. ISBN 0-07-913665-6
- ↑ Committee on Revising Science and Creationism: A View from the National Academy of Sciences, National Academy of Sciences and Institute of Medicine of the National Academies. 2008. Science, Evolution, and Creationism. National Academy of Sciences. Pristupljeno 6. siječnja 2008.
- ↑ Renne, P.R., Ludwig, K.R. and Karner, D.B. 1998. Progress and challenges in geochronology. Science Progress, 83, 107–121 [1]
- ↑ Noller, J.S., Sowers, J.M., and Lettis, W.R., 2000, Quaternary Geochronology: Methods and Applications, American Geophysical Union, AGU Reference Shelf 4.
- ↑ Colman, S. M., Pierce, K. L., and Birkeland, P. W., 1987, Suggested terminology for Quaternary dating methods: Quaternary Research, v. 28, p. 314–319
- ↑ Dickin, A. P. 1995. Radiogenic Isotope Geology. Cambridge, Cambridge University Press. ISBN 0-521-59891-5
- ↑ Faure, G. 1986. Principles of isotope geology. Cambridge, Cambridge University Press. ISBN 0-471-86412-9
- ↑ Faure, G., and Mensing, D. 2005. "Isotopes - Principles and applications". 3rd Edition. J. Wiley & Sons. ISBN 0-471-38437-2
- ↑ See:
- Dalrymple, G.B. 1991. The Age of the Earth. Stanford University Press. California. ISBN 0-8047-1569-6
- Newman, William L. 9. srpnja 2007. Age of the Earth. Publications Services, USGS. Pristupljeno 20. rujna 2007.
- Dalrymple, G. Brent. 2001. The age of the Earth in the twentieth century: a problem (mostly) solved. Geological Society, London, Special Publications. 190 (1): 205–221. Bibcode:2001GSLSP.190..205D. doi:10.1144/GSL.SP.2001.190.01.14. Pristupljeno 20. rujna 2007.
- Stassen, Chris. 10. rujna 2005. The Age of the Earth. TalkOrigins Archive. Pristupljeno 30. prosinca 2008.
- ↑ Bing Shen, Lin Dong,Shuhai Xiao, Michał Kowalewski The Avalon Explosion: Evolution of Ediacara Morphospace Science 4 January 2008: Vol. 319 no. 5859 pp. 81–84, DOI:10.1126/science.1150279
- ↑ Legendre, Serge. 1989. Les communautés de mammifères du Paléogène (Eocène supérieur et Oligocène) d'Europe occidentale : structures, milieux et évolution. F. Pfeil. München. str. 110. ISBN 978-3-923871-35-3
- ↑ Romer, A. S. 1933. Vertebrate Paleontology. University of Chicago Press, 3rd , 1966.
- ↑ therapsid (fossil reptile order) - Encyclopedia Britannica
- ↑ M. Laurin & R. R. Reisz. 1996. The osteology and relationships of Tetraceratops insignis, the oldest known therapsid. Journal of Vertebrate Paleontology 16(1): 95-102.
- ↑ J. Liu, B. Rubidge & J. Li, New basal synapsid supports Laurasian origin for therapsids, 2009, Acta Palaeontol. Pol., 54 (3): 393-400
- ↑ Dawkins R (2004) The Ancestor's Tale. "Query: Hominidae/Hylobatidae". Time Tree. 2009. Retrieved December 2013.
- ↑ Stringer, C.B. 1994. Evolution of Early Humans. The Cambridge Encyclopedia of Human Evolution. Cambridge University Press. Cambridge. str. 242. ISBN 978-0-521-32370-3 Nepoznati parametar
|editors=zanemaren (prijedlog zamjene:|editor=) (pomoć) Also ISBN 978-0-521-46786-5 (paperback) - ↑ McHenry, H. M. 2009. Human Evolution. Evolution: the first four billion years. The Belknap Press of Harvard University Press. Cambridge, Massachusetts. str. 265. ISBN 978-0-674-03175-3 Nepoznati parametar
|editors=zanemaren (prijedlog zamjene:|editor=) (pomoć) - ↑ g Out of Africa Revisited Provjerite vrijednost parametra
|url=(pomoć). Science. 308 (5724): 921 g. 13. svibnja 2005. doi:10.1126/science.308.5724.921g. Inačica izvorne stranice arhivirana 22. studenoga 2010. - ↑ Nature. 12. lipnja 2003. Access: Human evolution: Out of Ethiopia. Nature. Inačica izvorne stranice arhivirana 22. studenoga 2010. Pristupljeno 23. studenoga 2009.
- ↑ Origins of modern humans: multiregional or out of Africa?. ActionBioscience. Inačica izvorne stranice arhivirana 22. studenoga 2010. Pristupljeno 23. studenoga 2009.
- ↑ Klages, Arthur (2008) "Sahelanthropus tchadensis: An Examination of its Hominin Affinities and Possible Phylogenetic Placement," Totem: The University of Western Ontario Journal of Anthropology: Vol. 16: Iss. 1, Article 5. http://ir.lib.uwo.ca/totem/vol16/iss1/5
- ↑ Reynolds, Sally C; Gallagher, Andrew. 29. ožujka 2012. African Genesis: Perspectives on Hominin Evolution. ISBN 9781107019959
- ↑ Perlman, David. July 12, 2001. Fossils From Ethiopia May Be Earliest Human Ancestor. National Geographic News. Pristupljeno July 2009.
Another co-author is Tim D. White, a paleoanthropologist at UC-Berkeley who in 1994 discovered a pre-human fossil, named Ardipithecus ramidus, that was then the oldest known, at 4.4 million years.
Provjerite vrijednost datuma u parametru:|accessdate=(pomoć) - ↑ Johanson i Maitland 1981, str. 283–297 Pogreška u predlošku harvnb: ne postoji izvor s oznakom: CITEREFJohansonMaitland1981 (pomoć)
- ↑ Johanson, D.C. 2009. Lucy (Australopithecus afarensis). Evolution: The First Four Billion Years. The Belknap Press of Harvard University Press. Cambridge, Massachusetts. str. 693–697. ISBN 978-0-674-03175-3 Nepoznati parametar
|editors=zanemaren (prijedlog zamjene:|editor=) (pomoć) - ↑ Wood, B.A. 1994. Evolution of australopithecines. The Cambridge Encyclopedia of Human Evolution. Cambridge University Press. Cambridge, U.K.. ISBN 978-0-521-32370-3 Nepoznati parametar
|editors=zanemaren (prijedlog zamjene:|editor=) (pomoć) Also ISBN 0-521-46786-1 (paperback). - ↑ Dawkins, Richard. 2004. The Ancestor's Tale: A Pilgrimage To the Dawn of Life. Weidenfeld & Nicolson. London. str. 77. ISBN 0-297-82503-8
- ↑ Hazarika, Manji. 16–30 lipnja 2007. Homo erectus/ergaster and Out of Africa: Recent Developments in Paleoanthropology and Prehistoric Archaeology (PDF)
- ↑ Mammal Anatomy: An Illustrated Guide. Marshall Cavendish Corporation. 2010. ISBN 0-7614-7882-5
- ↑ Dawkins, Richard. 2009. 11. History Written All Over Us. The Greatest Show on Earth. Free Press. New York. ISBN 978-1-4165-9478-9. Pristupljeno 21. studenoga 2009.
- ↑ Kirk, Geoffrey; Raven, John; Schofield, John. 1984. The Presocratic Philosophers: A Critical History with a Selection of Texts treće izdanje izdanje. izdavač: The University of Chicago Press. Chicago. str. 100–142. ISBN 0-521-27455-9
- ↑ Kirk, Geoffrey; Raven, John; Schofield, John. 1984. The Presocratic Philosophers: A Critical History with a Selection of Texts treće izdanje izdanje. izdavač: The University of Chicago Press. Chicago. str. 280–321. ISBN 0-521-27455-9
- ↑ Bowler, Peter J. 2003. Evolution: the history of an idea. Berkeley, CA. str. 73.–75
- ↑ Carl Linnaeus - berkeley.edu. Pristupljeno 11. veljače 2012.
- ↑ Gould, S.J. 2002. The Structure of Evolutionary Theory. izdavač: Belknap Press (Harvard University Press). Cambridge. ISBN 0-674-00613-5
- ↑ Wright, S. Evolution and the Genetics of Populations, Volume 1: Genetic and Biometric Foundations. izdavač: University Of Chicago Press. str. str. 480. ISBN 0-226-91038-5
- ↑ Sturm RA, Frudakis TN. 2004. Eye colour: portals into pigmentation genes and ancestry. Trends Genet. 20 (8): 327–32. doi:10.1016/j.tig.2004.06.010. PMID 15262401
- ↑ a b Pearson H. 2006. Genetics: what is a gene?. Nature. 441 (7092): 398–401. Bibcode:2006Natur.441..398P. doi:10.1038/441398a. PMID 16724031
- ↑ Visscher PM, Hill WG, Wray NR. 2008. Heritability in the genomics era—concepts and misconceptions. Nature Reviews Genetics. 9 (4): 255–66. doi:10.1038/nrg2322. PMID 18319743CS1 održavanje: više imena: authors list (link)
- ↑ Oetting WS, Brilliant MH, King RA. 1996. The clinical spectrum of albinism in humans. Molecular medicine today. 2 (8): 330–5. doi:10.1016/1357-4310(96)81798-9. PMID 8796918CS1 održavanje: više imena: authors list (link)
- ↑ Futuyma, Douglas J. 2005. Evolution. Sinauer Associates, Inc. Sunderland, Massachusetts. ISBN 0-87893-187-2
- ↑ Phillips PC. 2008. Epistasis—the essential role of gene interactions in the structure and evolution of genetic systems. Nature Reviews Genetics. 9 (11): 855–67. doi:10.1038/nrg2452. PMC 2689140. PMID 18852697
- ↑ a b Wu R, Lin M. 2006. Functional mapping – how to map and study the genetic architecture of dynamic complex traits. Nature Reviews Genetics. 7 (3): 229–37. doi:10.1038/nrg1804. PMID 16485021
- ↑ Jablonka, E.; Raz, G. 2009. Transgenerational epigenetic inheritance: Prevalence, mechanisms and implications for the study of heredity and evolution. The Quarterly Review of Biology. 84 (2): 131–176. doi:10.1086/598822. PMID 19606595
- ↑ Bossdorf, O.; Arcuri, D.; Richards, C. L.; Pigliucci, M. 2010. Experimental alteration of DNA methylation affects the phenotypic plasticity of ecologically relevant traits in Arabidopsis thaliana. Evolutionary Ecology. 24 (3): 541–553. doi:10.1007/s10682-010-9372-7
- ↑ Jablonka, E.; Lamb, M. 2005. Evolution in four dimensions: Genetic, epigenetic, behavioural and symbolic. MIT Press. ISBN 0-262-10107-6
- ↑ Wassersug, J. D., and R. J. Wassersug, 1986. Fitness fallacies. Natural History 3:34-37.
- ↑ Maynard-Smith, J. (1989) Evolutionary Genetics ISBN 0-19-854215-1
- ↑ Harwood AJ; Harwood, J. 1998. Factors affecting levels of genetic diversity in natural populations. Philosophical Transactions of the Royal Society B. 353 (1366): 177–86. doi:10.1098/rstb.1998.0200. PMC 1692205. PMID 9533122
- ↑ Ewens W.J. 2004. Mathematical Population Genetics (2nd Edition). Springer-Verlag, New York. ISBN 0-387-20191-2
- ↑ Butlin RK, Tregenza T. 1998. Levels of genetic polymorphism: marker loci versus quantitative traits. Philosophical Transactions of the Royal Society B. 353 (1366): 187–98. doi:10.1098/rstb.1998.0201. PMC 1692210. PMID 9533123
- ↑ Wetterbom A, Sevov M, Cavelier L, Bergström TF. 2006. Comparative genomic analysis of human and chimpanzee indicates a key role for indels in primate evolution. J. Mol. Evol. 63 (5): 682–90. doi:10.1007/s00239-006-0045-7. PMID 17075697CS1 održavanje: više imena: authors list (link)
- ↑ Sawyer SA, Parsch J, Zhang Z, Hartl DL. 2007. Prevalence of positive selection among nearly neutral amino acid replacements in Drosophila. Proc. Natl. Acad. Sci. U.S.A. 104 (16): 6504–10. Bibcode:2007PNAS..104.6504S. doi:10.1073/pnas.0701572104. PMC 1871816. PMID 17409186CS1 održavanje: više imena: authors list (link)
- ↑ Hastings, P J; Lupski, JR; Rosenberg, SM; Ira, G. 2009. Mechanisms of change in gene copy number. Nature Reviews Genetics. 10 (8): 551–564. doi:10.1038/nrg2593. PMC 2864001. PMID 19597530
- ↑ Sean B. Carroll; Jennifer K. Grenier; Scott D. Weatherbee. 2005. From DNA to Diversity: Molecular Genetics and the Evolution of Animal Design. Second Edition. Blackwell Publishing. Oxford. ISBN 1-4051-1950-0CS1 održavanje: više imena: authors list (link)
- ↑ Harrison P, Gerstein M. 2002. Studying genomes through the aeons: protein families, pseudogenes and proteome evolution. J Mol Biol. 318 (5): 1155–74. doi:10.1016/S0022-2836(02)00109-2. PMID 12083509
- ↑ Bowmaker JK. 1998. Evolution of colour vision in vertebrates. Eye (London, England). 12 (Pt 3b): 541–7. doi:10.1038/eye.1998.143. PMID 9775215
- ↑ Gregory TR, Hebert PD. 1999. The modulation of DNA content: proximate causes and ultimate consequences. Genome Res. 9 (4): 317–24. doi:10.1101/gr.9.4.317. PMID 10207154
- ↑ Hurles M. 2004. Gene duplication: the genomic trade in spare parts. PLoS Biol. 2 (7): E206. doi:10.1371/journal.pbio.0020206. PMC 449868. PMID 15252449
- ↑ Liu N, Okamura K, Tyler DM. 2008. The evolution and functional diversification of animal microRNA genes. Cell Res. 18 (10): 985–96. doi:10.1038/cr.2008.278. PMC 2712117. PMID 18711447CS1 održavanje: više imena: authors list (link)
- ↑ Siepel A. 2009. Darwinian alchemy: Human genes from noncoding DNA. Genome Res. 19 (10): 1693–5. doi:10.1101/gr.098376.109. PMC 2765273. PMID 19797681
- ↑ Orengo CA, Thornton JM. 2005. Protein families and their evolution-a structural perspective. Annu. Rev. Biochem. 74 (1): 867–900. doi:10.1146/annurev.biochem.74.082803.133029. PMID 15954844
- ↑ Long M, Betrán E, Thornton K, Wang W. 2003. The origin of new genes: glimpses from the young and old. Nature Reviews Genetics. 4 (11): 865–75. doi:10.1038/nrg1204. PMID 14634634CS1 održavanje: više imena: authors list (link)
- ↑ Wang M, Caetano-Anollés G. 2009. The evolutionary mechanics of domain organisation in proteomes and the rise of modularity in the protein world. Structure. 17 (1): 66–78. doi:10.1016/j.str.2008.11.008. PMID 19141283
- ↑ Weissman KJ, Müller R. 2008. Protein-protein interactions in multienzyme megasynthetases. Chembiochem. 9 (6): 826–48. doi:10.1002/cbic.200700751. PMID 18357594
- ↑ Radding C. 1982. Homologous pairing and strand exchange in genetic recombination. Annu. Rev. Genet. 16 (1): 405–37. doi:10.1146/annurev.ge.16.120182.002201. PMID 6297377
- ↑ Agrawal AF. 2006. Evolution of sex: why do organisms shuffle their genotypes?. Curr. Biol. 16 (17): R696–704. doi:10.1016/j.cub.2006.07.063. PMID 16950096
- ↑ Peters AD, Otto SP. 2003. Liberating genetic variance through sex. BioEssays. 25 (6): 533–7. doi:10.1002/bies.10291. PMID 12766942
- ↑ Goddard MR, Godfray HC, Burt A. 2005. Sex increases the efficacy of natural selection in experimental yeast populations. Nature. 434 (7033): 636–40. Bibcode:2005Natur.434..636G. doi:10.1038/nature03405. PMID 15800622CS1 održavanje: više imena: authors list (link)
- ↑ Morjan C, Rieseberg L. 2004. How species evolve collectively: implications of gene flow and selection for the spread of advantageous alleles. Mol. Ecol. 13 (6): 1341–56. doi:10.1111/j.1365-294X.2004.02164.x. PMC 2600545. PMID 15140081
- ↑ Boucher Y, Douady CJ, Papke RT, Walsh DA, Boudreau ME, Nesbo CL, Case RJ, Doolittle WF. 2003. Lateral gene transfer and the origins of prokaryotic groups. Annu Rev Genet. 37 (1): 283–328. doi:10.1146/annurev.genet.37.050503.084247. PMID 14616063CS1 održavanje: više imena: authors list (link)
- ↑ Walsh T. 2006. Combinatorial genetic evolution of multiresistance. Current Opinion in Microbiology. 9 (5): 476–82. doi:10.1016/j.mib.2006.08.009. PMID 16942901
- ↑ Kondo N, Nikoh N, Ijichi N, Shimada M, Fukatsu T. 2002. Genome fragment of Wolbachia endosymbiont transferred to X chromosome of host insect. Proc. Natl. Acad. Sci. U.S.A. 99 (22): 14280–5. Bibcode:2002PNAS...9914280K. doi:10.1073/pnas.222228199. PMC 137875. PMID 12386340CS1 održavanje: više imena: authors list (link)
- ↑ Sprague G. 1991. Genetic exchange between kingdoms. Current Opinion in Genetics & Development. 1 (4): 530–3. doi:10.1016/S0959-437X(05)80203-5. PMID 1822285
- ↑ Gladyshev EA, Meselson M, Arkhipova IR. 2008. Massive horizontal gene transfer in bdelloid rotifers. Science. 320 (5880): 1210–3. Bibcode:2008Sci...320.1210G. doi:10.1126/science.1156407. PMID 18511688CS1 održavanje: više imena: authors list (link)
- ↑ Baldo A, McClure M. 1. rujna 1999. Evolution and horizontal transfer of dUTPase-encoding genes in viruses and their hosts. J. Virol. 73 (9): 7710–21. PMC 104298. PMID 10438861
- ↑ River, M. C. and Lake, J. A. 2004. The ring of life provides evidence for a genome fusion origin of eukaryotes. Nature. 431 (9): 152–5. Bibcode:2004Natur.431..152R. doi:10.1038/nature02848. PMID 15356622CS1 održavanje: više imena: authors list (link)
- ↑ Scott EC, Matzke NJ. 2007. Biological design in science classrooms. Proc. Natl. Acad. Sci. U.S.A. 104 (suppl_1): str. 8669.–8676. Bibcode:2007PNAS..104.8669S. doi:10.1073/pnas.0701505104. PMC 1876445. PMID 17494747
- ↑ Hendry AP, Kinnison MT. 2001. An introduction to microevolution: rate, pattern, process. Genetica. 112–113: str. 1.–8. doi:10.1023/A:1013368628607. PMID 11838760
- ↑ Leroi AM (2000.). "The scale independence of evolution". Evol. Dev. 2 (2): str. 67.–77.
- ↑ Gould SJ. 1994. Tempo and mode in the macroevolutionary reconstruction of Darwinism. Proc. Natl. Acad. Sci. U.S.A. 91 (15): str. 6764.–6771. Bibcode:1994PNAS...91.6764G. doi:10.1073/pnas.91.15.6764. PMC 44281. PMID 8041695
- ↑ Jablonski, D. 2000. Micro- and macroevolution: scale and hierarchy in evolutionary biology and paleobiology. Paleobiology. 26 (sp4): str. 15.–52. doi:10.1666/0094-8373(2000)26[15:MAMSAH]2.0.CO;2